Фитоцен микрофитобентоса представлен преимущественно бентосными и перифитонными таксонами диатомей, нитчатыми и зооглейными формами синеэеленых водорослей и симбиотическими зооксантеллами фораминифер (Plante-Cuny, 1973; Gribb, 1973; Sournia, 1976, 1977; Miller et al., 1977). В числе основных родов диатомовых водорослей, населяющих донные осадки рифа, такие, как Navicula, Nitzschia, Surirella, Achanther, Cocconeis, Amphora, Diploneis, Gyrosigma, Bacillaria, Mastogloia,Rhoicosigma, Donkinia, Coscinodiscus. При этом одни их виды (Cocconeis, Achanthes) прикрепляются к песчинкам, а другие движутся между ними (Nitzschia, Navicula, Gyrosigma). Их численность в коралловом песке достигает иногда 5-7·105 кл/г.
В числе массовых видов синезеленых водорослей, обитающих в составе микрофитобентоса, доминируют представители родов Schizotrix, Calotrix, Rivularia, Oscilla-toria, Chroococcus, Lyngbia. Синсзеленые водоросли часто образуют сплошной слизистый покров на мелком обломочном материале и на крупном песке в зонах его аккумуляции, особенно в затишных участках. Диатомеи также образуют иногда сплошной покров зеленовато-бурого цвета в затишных участках дна лагун. На мелководных участках патч-рифов, флета и прорезающих его каналов в крупнозернистых коралловый песках в большом количестве обитают крупные фораминиферы, которые содержат симбиотические водоросли-зооксантеллы. На таких участках дна именно они, а не свободно живущие микроводоросли производят преобладающую часть автотрофной продукции (Plante-Cuny, 1973; Sournia, 1976). Содержание хлорофилла "а" в верхнем слое кораллового песка в среднем близко к 1,5-5 мкг/г (пределы колебаний от 0,5 до 30 мкг/г, Plante-Cuny, 1973; Sournia, 1976; Пропп и др., 1983; Burns et аl., 1984). Биомасса водорослей в коралловых песках, согласно нашим определениям, составляет в среднем 1-2% от общего органического вещества и выражается величинами 0,4-1,3 мг сырого веса на 1г (Sorokin, 1973, 1978а). Аналогичные величины дает расчет биомассы водорослей: исходя из среднего содержания хлорофилла "а" в коралловых песках 3-5 мкг/г и среднего соотношения "сырая биомасса водорослей" хлорофилла "а", равного 150:1 (отношение углерод биомассы : хлорофилл 10 : 1) . Суточные коэффициенты удельной продукции водорослей (Р/В) при таких значениях их биомассы выражаются величинами 0,1—0,4. Биомасса водорослей в составе микрофитобентоса в среднем в 1,5—3 раза ниже биомассы бактерий. Лишь в "регенеративном" (детритном) осадке биомасса водорослей, составлявшая 24,5 мг/г, оказалась выше биомассы бактерий.
Продукция фотосинтеза фитобентоса песков, загрязняемых рифов в заливе Канеохе оказалась относительно низкой (см. табл. 7.5). В расчете на 1 м2 она составляла 0,1-0,2 г С/м2 в сутки. В коралловый песках этого залива преобладали деструкционные процессы. Дыхание микрофлоры превышало в них в 3-10 раз фотосинтез фитобентоса, составляя 0,7-1,9 г С/м2 в сутки. В донных осадках атолла Маджуро, не подверженный антропогенному загрязнению, фотосинтез фитобентоса был в среднем в 2-5 раз выше, чем в донных осадках загрязняемого залива Канеохе, и существенно превышал бактериальную продукцию. В расчете на 1м2 суточная продукция фотосинтеза микрофитобентоса песков на большинстве исследованных участков была выше 0,3 г С/м2, а на отдельный из них достигала 1,5-2,2 г С/м2. Однако наибольшая ее величина была найдена в леске прибрежного пляжа у верхней границы прилива (11,7 г С/м2 в сутки), что было соизмеримо с продукцией поля макрофитов или зарослей кораллов.
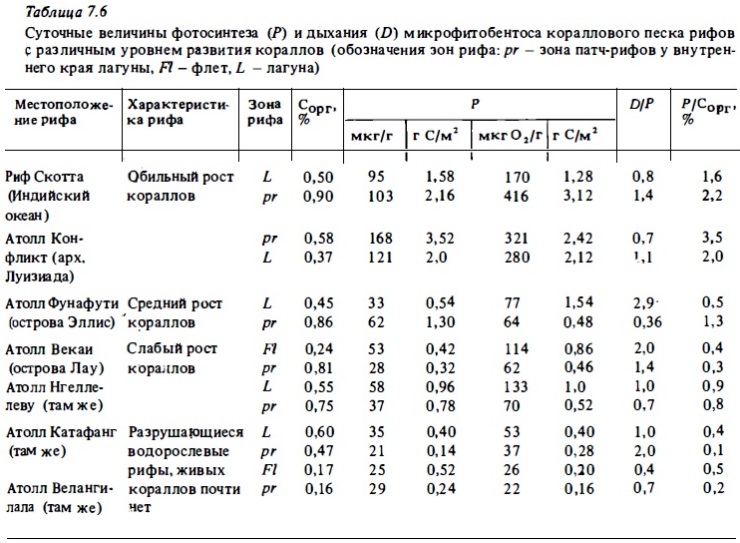
Интенсивность дыхания и фотосинтеза микрофитобентоса в коралловых. песках атоллов зависит от степени заселенности окружающих рифов кораллами. На процветающих рифах с обильным ростом кораллов эти показатели метаболизма сообществ донной микрофлоры были в 5-10 раз выше, чем в донных осадках водорослевых рифов, находившихся в угнетенном состоянии (Сорокин, 1980; Sorokin, 1981b). Суточный фотосинтез в коралловых песках на процветающих рифах достигал 3,5 г С/м2, дыхание - 4,16 г С/м2 в сутки. В расчете на 1 г сухого веса фотосинтез в коралловом песке таких рифов составлял 100-160 мкг С/г в сутки, что близко к суточному фотосинтезу живых кораллов в расчете на вес их колоний. Дыхание на процветающих рифах, как правило, превышает фотосинтез фитобентоса в 1,5-3 раза. Это свидетельствует в пользу того, что в донные осадки таких рифов в большом количестве поступает усвояемое органическое вещество, в основном в виде слизи, выделяемой кораллами. Приток органического вещества. в донные осадки стимулирует как; процессы деструкции, так и развитие фитобентоса в верхнем слое осадков, поскольку в условиях интенсивной деструкции усиливается и поставка солевых биогенов сообществам обитающего в них фитобентоса. В донных осадках рифов со слабым ростом кораллов поставка органического вещества (а вместе с ним и биогенов) снижается, что приводит к снижению интенсивности фотосинтеза и дыхания фитобентоса. При этом дыхание снижается в большей степени, чем фотосинтез, который в донных осадках таких рифов обычно превышает дыхание в 1,5—5 раз (табл. 7.6).
Высокий уровень фотосинтеза и дыхания отмечен в донных осадках коралловых рифов Центрального Вьетнама, которые характеризуются интенсивным поступлением органического вещества за счет терригенного стока. Коралловые пески приконтинентальных рифов часто заносятся терригенными наилками. Донные осадки мелководных участков характеризуются эдесь весьма интенсивным метаболизмом и высокой продукцией фотосинтеза обитающего в них микрофитобентоса. Суточный фотосинтез фитобентоса на большинстве обследованных рифов был выше 0,4 г С/м2, превышая в отдельных случаях 1 г С/м2. При этом на большей части станций в мелководных участках дна фотосинтез в суточном балансе был выше дыхания. Интенсивность дыхания составляла в среднем 0,4-1 г С/м2. На приглубых участках дна, глубже 5 м, фотосинтез снижался до 0,2-0,6 г С/м2. Дыхание на приглубых участках дна в большинстве случаев превышало фотосинтез в суточном балансе. Наибольшие величины дыхания (до 6 г О2/м2 в сутки) были зарегистрированы в карбонатных илах, которые отлагаются в приглубых участках бухт. При такой интенсивности потребления кислорода может возникать его дефицит в придонном слое, что способствует возникновению сульфатредукции. Сульфатредукция широко распространена в заиленных коралловых песках прибрежных рифов Вьетнама, где слой черных. от выпадения гидротроилита осадков часто можно обнаружить на глубине 1-2 см под поверхностью дна.
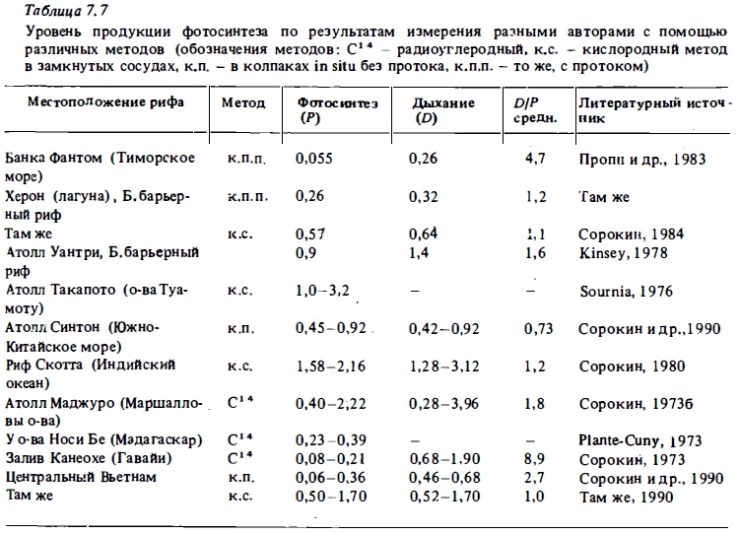
Сведения об интенсивности фотосинтеза и дыхания микрофитобентоса донных осадков рифов, полученные различными авторами, обобщены в табл. 7.7. Они показывают, что в целом применение разных методов для ее определения дает достаточно сопоставимые результаты. Суточный фотосинтез коралловых песков на мелководьях близок в среднем к 0,3-1 г С/м2. Дыхание в большинстве случаев превышает фотосинтез, что особенно четко выражено в зонах обильного роста кораллов или терригенного стока органики, когда обеспечивается дополнительно к фотосинтезу поставка усвояемого органического вещества гетеротрофной микрофлоре донных осадков.