Бактериальное население перифитонных обрастаний представлено преимущественно прикрепляющимися нитевидными бактериями типа флексибактерий и бактерий родов Cladporix, Leptotrix и Crenotrix. Особенно многочисленны в перифитснных обрастаниях флексибактерии, именуемые и скользящими бактериями. Их нити постоянно активно изгибаются, вследствие чего увеличивается их контакт клеток с окружающей водной средой. Многочисленны также в составе перифитона одиночные бактерии, обладающие выростами для прикрепления к поверхности клеток перифитонных водорослей, такие, как каулобактерии и простекобактерии.
В состав водорослей перифитона входят мелкие талломные макрофиты с размером талломов 0,5—1,5см, корковые кораллины, нитчатки, микроводоросли с размерами от 2 до 100 км. В составе талломных макрофитов перифитона чаще других встречаются представители зеленых водорослей из родов Enteromorpha, Caulerpa, Chaetomorpha, Acetabularia, Bryopsys, бурых водорослей Ectocarpus, Giffordia, Sphacelaria и красных водорослей Gelidiella, Polisiphonia, Chondria, Champia, Heterosiphonia, Laurencia, Hypnea, Plerocladia, Wurdemannia. Нитчатки, обильно заселяющие мертвые кораллы и обломочный материал, представлены в основном зелеными водорослями Cladophora, Cladopho-ropsis, Trichosolen и синезелеными нитчатыми водорослями, которые часто доминируют в составе фитоцена перифитонных обрастаний, образуя на них сплошной покров. Среди них массовыми являются представители родов Calotrix, Schizotrix, Kyritrix, Oscillatoria, Mastigocoleus.
Важное значение в формировании фитоцена перифитона имеют такие представители таких родов синезеленых водослей, как Spirulina, Lyngbia, Nostoc, Rivularia. Синезеленые водоросли Nostoc, Rulfsia, Entophysalia образуют смолоподобные темно-зеленые или черно-коричневые слизистые налеты на скальном флете и на поверхности мертвых кораллов. Некоторые из широко распространенных на рифах зеленых и синезеленых нитчатых водорослей способны "сверлить" известняк. Они проникают в толщу приповерхностных слоев скального флета, в колонии мертвых и живых кораллов и плотно их заселяют. В числе таких водорослей зеленая водоросль Ostreobium reineckei и синезеленые водоросли Entophysalia deusta и Mastigocoleus testarum (Odum, Odum, 1955; Gribb, 1973; Kobluk, Risk, 1977).
В составе перифитона в массовом количестве встречаются красные корковые известковые водоросли кораллины. Они часто образуют сплошные корки на колониях отмерших ветвистых кораллов и на поверхности скального флета. Особенно они доминируют в составе перифитонных обрастаний на затененных их поверхностях, покрывая нижние части колоний отмерших кораллов, стенки расщелин и пещер скального флета. Среди них в составе перифитона обычны представители родов Porolithon, Jania, Peys-sonelia, Litothamnion, Lithophyllum, Neogoniolithon. Наконец в составе перифитона обитает множество разнообразным микроводорослей размерами до 5 мкм и менее. Среди них различные донные и перифитонные диатомеи, которые представлены такими их родами, как Camylodiscus, Podocystis, Triceratium, Navicula, Nitzschia. Некоторые из перечисленным видов диатомей обитают и на живых кораллах (Montgomery et аl., 1977). В составе микроводорослей перифитона, численность которым достигает 105 кл/см2, представители мелких синезеленьм водорослей из порядка Nostocales. В состав сообществ перифитона входят также организмы микробентоса. Они представлены прежде всего инфузориями, мелкими фораминиферами и мелкими нематодами. Биомасса бактерий и микроводорослей в составе перифитона составляет от долей шллиграммов до нескольких миллиграммов на квадратный сантиметр (0,4-4,2 мг/см2), или 5-15% от общего органического вещества обрастаемых им отростков ветвистых колоний мертвых кораллов (табл. 7.8, 7,9). Соотношение между биомассой водорослей и биомассой бактерий варьировало от 3 на загрязняемом участке залива (станция 1) до 200 на чистом участке внешнего рифа. Соответственно в первом случае биомасса и суточная продукция бактериального населения перифитона были почти на порядок выше, чем в последнем. Эти данные показывают, что в условиях загрязнения развитие перифитонных водорослей угнетается и их место занимают бактерии. В чистых участках соотношение между ними зависит в большей степени от механизмов, регулирующих отношения между автотрофами и гетеротрофами в сообществе.
Общая численность бактерий в составе перифитонных обрастаний, как правило, приближается к 109 кл/см2 (см. табл. 7.8, 7.9), а в расчете на 1 г соскоба составляет 2-6·109 кл (табл. 7.10). При этом в отличие от микрофлоры микрофитобентоса обилие микрофлоры в перифитонных обрастаниях разных рифов практически не зависит от заселенности их живыми кораллами (табл. 7.11). Микрофлора перифитона одинаково обильна на живом процветающем рифе Скотта, и на голом флете разрушенного атолла Фунафути, и на угнетенныи окаймляюших рифах Новых Гебрид и архипелага Лау, где живые кораллы практически отсутствуют. Аналогичным образом ведет себя и водорослевый компонент перифитона, если судить по интенсивности его фотосинтеза, а также по общей интенсивности метаболизма перифитона. Эти данные показывают, что сообщества перифитона рифов получают органическое вещество и биогены из внешнего по отношению к экосистеме самого рифа источника, каким являются проходящие над рифом воды океана. Эти вода даже в олиготрофных районах содержат 0,1—0,2 мкг ат./л доступного для бактерий перифитона органического фосфора и 0,2-0,4 мг С/л усвояемого для бактерий перифитона органического вещества. Описанные свойства перифитонньи обрастаний имеют первостепенное значение для функционирования экосистемы рифа, поскольку они способствуют обеспечению положительного баланса биогенов и органического вещества при ее взаимодействии с окружающим океаном.
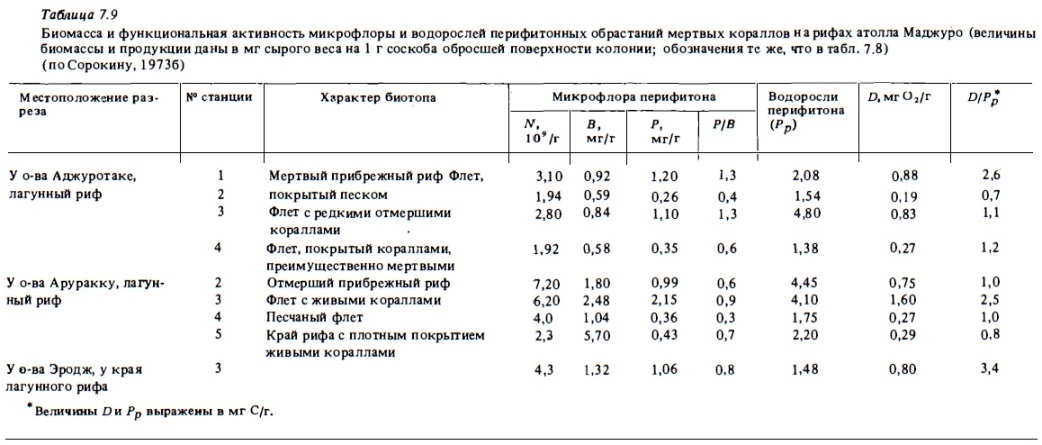
Что касается интенсивности метаболизма перифитонных обрастаний, то уже его первые анализы, выполненные нами на гавайских рифах и на атолле Маджуро (см. табл.7.8, 7.9), показали, что по интенсивности фотосинтеза и дьиания их можно считать в числе наиболее активных сообществ экосистемы рифа. Коэффициенты удельной продукции микрофлоры рифа (Р/В) оказались значительно (в 2-3 раза) выше таковых у микрофлоры донных осадков (см. табл. 7.2). Удельная продукция водорослей перифитона (0,3-0,6 за сутки) также оказалась весьма высокой и соизмеримой с таковой у фитопланктона рифа. Суточная продукция фотосинтеза в расчете на 1 см2 поверхности колоний мертвых кораллов на внешних гавайских рифах в заливе Канеохе достигала 1,1 мг С/м2 в сутки, что в расчете на 1 м2 в зависимости от степени покрытия ими дна может дать до 10 г С/м2 органической продукции за сутки. В загрязненном участке залива Канеохе (см. табл. 7.5) развитие перифитона было ослабленным. Однако и здесь суточный фотосинтез превышал 0,3 мг С/м2 в сутки. На загрязняемом участке дыхание перифитона бьшо существенно выше фотосинтеза. Это указывает на преобладание гетеротрофных деструкционных процессов в сообществах перифитона загрязняемой зоны рифа. На чистом атоме Маджуро перифитонные обрастания характеризовались очень высокими показателями биомассы входящей в их состав микрофлоры (1-5 мг/г) и высокой ее продукцией (0,3-2 мг/г в сутки) при величине коэффициентов Р/В 0,6-1,3. Интенсивность фотосинтеза водорослей перифитонных обрастаний была здесь также очень велика и достигала 2-4 мг сырой биомассы на 1 г соскоба перифитона или 120-250 мгк С/г в сутки. При этом фотосинтез перифитонных обрастаний в зоне лагунных рифов этого атолла был в большинстве случаев сбалансирован с дыханием. Результаты измерений, выполненных в рифах Вьетнама (см. табл. 7.10), показали, что в перифитонных обрастаниях приконтинентальных рифов, получающих значительный приток органического вещества с суши, микрофлора намного богаче, чем на рифах в открытых районах Южно-Китайского моря. Суточная продукция фотосинтеза перифитона превышала в большинстве случаев 1 г С/м2, а на атолле Синтон достигала 3 г С/м2 в сутки. При этом фотосинтез перифитона в суточном балансе был, как правило, в 2-6 раз выше дыхания. Лишь в бухте Ванфонг, подверженной загрязнению, дыхание было выше фотосинтеза.
Основные сведения об интенсивности автотрофной продукции и гетеротрофного метаболизма перифитонных обрастаний разных рифов обобщены в табл. 7.12. Они показывают, что в среднем продукция и метаболизм перифитонных обрастаний в расчете на 1 м2 площади рифа в 1,5-2 раза выше, чем у микрофитобентоса донных осадков. По средним данным, оба эти показателя в определенной мере сбалансированы и выражаются достаточно высокими величинами: 1-2 г С/м2 в сутки. Учитывая необычайно широкое развитие перифитонных обрастаний на рифах, можно с уверенностью утверждать, что сообщества перифитона следует отнести по их значимости к ключевым функциональным компонентам экосистемы рифа наряду с сообществами самих кораллов. Наглядным подтверждением такого вывода могут служить результаты опытов, доказавших соизмеримость интенсивности метаболизма живых кораллов и отмерших участков их колоний, покрытых перифитоном в расчете на вес колонии (табл. 7.13).