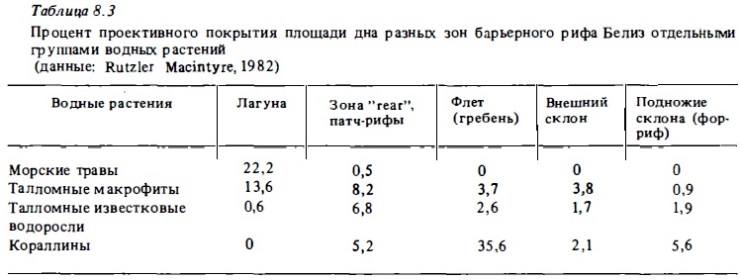
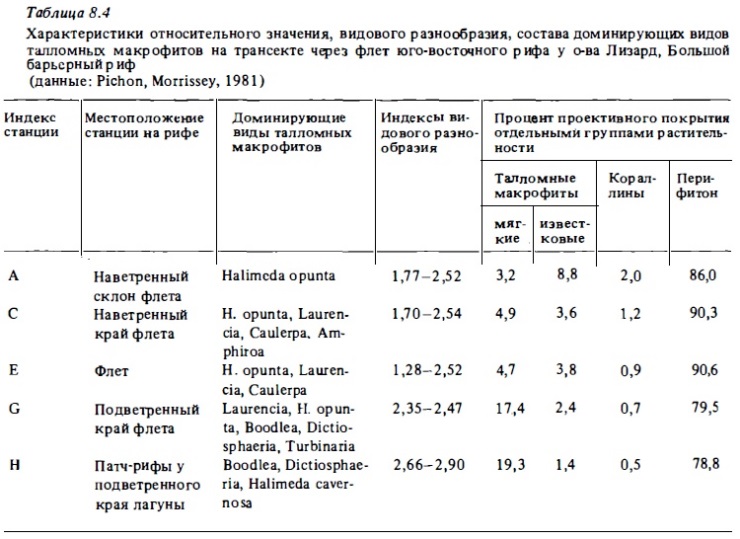
В качестве примера зонального распределения массовый видов водорослей на рис. 8.2 представлены соответствующие сведения для разреза через барьерный риф у Ямайки. Как видно из этого рисунка, большинство массовых видов даже в пределах одного рода специализированы в отношении предпочитаемых биотопов. Наиболее оппортунистическим среди массовых видов макрофитов рифа оказалась ветвящаяся кораллина из рода Peyssonnelia, которая была слабо представлена лишь в лагуне, обитая в ней только на поверхности колоний мертвых кораллов и крупного обломочного материала. Наибольшим видовым разнообразием характеризовались на этом разрезе зона каменистой литорали у берега острова, зона патч-рифов у тыльной стороны наветренного флета и в несколько меньшей степени приглубые участки внешнего склона, где волновой стресс снижен.
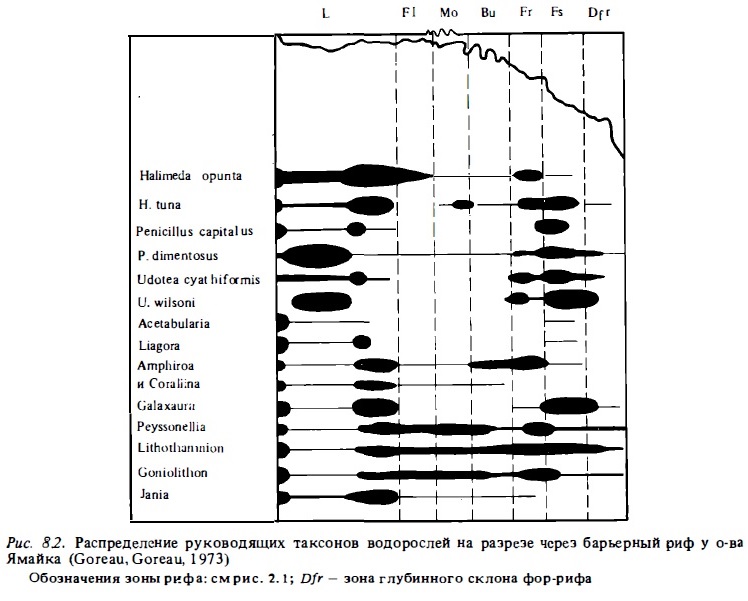
Представление о зональном распределении отдельных групп водной растительности на рифах разного типа дают табл. 8.3, 8.4. В них приводятся величины проективного покрытия площади дна в разных зонах рифов отдельными группами растений. На барьерном рифе Белиз, для которого характерен полупогруженный флет со значительным покрытием флета и зоны патч-рифов живыми кораллами, макрофиты занимают меньше половины площади. В лагуне этого рифа преобладает ассоциация морской травы и макрофитов. В зоне флета на его гребне в условиях волнобоя, сильной радиации и периодической осушки растительная ассоциация представлена кораллинами, которые в наибольшей степени приспособлены к таким условиям.
На другом рифе, у о-ва Лизард, где флет подвергается более значительному воздействию волнения, основные площади скальных поверхностей флета покрыты перифитоном, в состав которого входят нитчатые и мелкоталломные водоросли и кораллины. На флете перифитон занимает до 90% всей его поверхности. Для растительных ассоциаций макрофитов, обитающих на флете в зоне волнобоя, характерно доминирование в их составе известковой зеленой водоросли Halimeda opunta с другими зелеными (Caulerpa) и красными (Laurencia, Amphiroa) водорослями, которые приспособлены к жизни в прибойной зоне. На подветренной стороне флета, вне волнового стресса, флора меняется. В ее составе появляются Boodlea, Dictyosphaeria и Halimeda cavernosa, предпочитающие более спокойные воды (Pichon, Morrissey, 1981).
Чтобы иметь представление о зональном распределении и составе растительных ассоциаций на рифах в разных географических зонах, ниже приводятся наиболее характерные конкретные примеры такого распределения на ряде рифов.
На окаймляющем рифе у о-ва Кюрасао зональное распределение водной растительности выглядит следующим образом. В верхней скальной литорали доминируют обрастания слизистых и нитчатых форм синезеленых водорослей. На нижней литорали доминируют Laurencia и Sargassum. На флете на разных его участках преобладают плотные "ковровые" заросли мелкоталломных макрофитов: Sphacelaria tribuloides, Pterocladia americana, Heterosiphonia tenella, Polysiphonia subtilissima, Wurdemannia miniata. Ближе к краю флета в нем преобладают корковые кораллины Porolithon pachydermum, Lithophyllum daedaleum и L. intermedium. На склоне, покрытом обломочным материалом, доминируют кораллины Hydrolithon boerefesenii и Archaeolitho-thamnium dinotum. Нижняя граница распределения кораллин лежит на глубине около 90 м, где освещенность составляла всего около 0,2% от приповерхностной (Wanders, 1976; Van den Hoeck et аl., 1978).
На атолле Диего Гарсия в зоне прибрежных патч-рифов обитают Cladophora, Crypto-nemia, Valonia, Botriocladia, Caulerpa, Avrainvillea, Bryopsis, Codium, Galaxaura, Schizotrix. Лагунные патч-рифы заселены макрофитами Valonia, Dictiosphaeria, Caulerpa, Autabularia, Cymodocea. На талломах последней в массе развиваются эпифиты Sphacelaria furcigera, Amphiroa aragillissima, Jania tenella, Champia parvula. На.песчаном дне лагуны с глубиной 10 м преобладает Halimeda. На колониях мертвых кораллов и на крупных обломках и вокруг пил на дне лагуны обитают Sargassum, Turbinaria, Padina, Jania, Galaxaura, Dictyota. Флет внешнего рифа, припорошенный песком, занят плотным растительным покровом, который состоит из нитчатых синезеленых и зеленых водорослей, мелкоталломньк макрофитов Dictiosphaeria, Boodlea, Gelidiella, Laurencia, Heterosiphonia, а также кораллины Jania. На отдельных участках скального флета образуются большие темные слизистые натеки, которые формируют синезеленые водоросли Rivularia polyotis, Microcoleus lyngbiaceus, Schizotrix calcicola, а также корковые кораллины. В трещинах флета обитает Halimeda, а на стенках кораллов прорезывающих флет, — нитчатка Cladophora. На прибойной части у края флета доминируют кораллины, а также макрофиты с мелкими прозрачными талломами — Microdictyon, Boodlea, Turbinaria, Botriocladia, Pocokiella, Laurencia и нитчатые синезеленые водоросли Schizotrix. Многие из них обитают в расщелинах края флета (Rhyne, 1971).
Особую роль в формировании фитоцена наветренного флета играют кораллины Porolithon, Neogoniolithon, Sporolithon, Lithophyllum и Hydrolithon. Их массивные, слабо расчлененные или бугрообразные колонии накрепко прирастают к скальному основанию флета и выносят волновой стресс. При этом отдельные виды кораллин занимают обычно разные экологические ниши, имея соответствующий спектр экологических адаптаций (Littler, 1972; Doty, 1974). Соответственно разные виды кораллин характеризуются своим типом распределения на разрезах через риф (Van den Hoeck et аl., 1978). Так, на гавайском окаймляющем рифе, где кораллины занимают более 30—50% всей поверхности наветренного флета, каждый из четырех доминирующих их видов: Porolithon onkoides, Р. gardineri, Sporolithon erithaeum и Hydrolithon reingoldii имеет свою зону доминирования. Примечательным среди этих видов является Р. onkoides, который следует признать наиболее оппортунистическим видом среди кораллин, обитающих на рифах. Этот вид выдерживает самую сильную прямую солнечную радиацию, длительную осушку. Он образует мощные бугровидные колонии, которые прочно прирастают к скальному субстрату и прекрасно выдерживают волно-бой. Развиваясь в зоне волнобоя у края флета, Р. onkoides избегает выедания рыбами (Adey, 1975). Благодаря описанным свойствам Р. onkoides, как правило, доминирует среди кораллин на наветренных краях флета и "водорослевого вала". Он выступает как главный рифостроитель в этой зоне рифа, принимающий на себя основной волновой стресс (см. рис. 8.1). Два других массовых вида кораллин Sporolithon и Hydrolithon характеризуются тенелюбивостью и не столь устойчивы к волнобою, как Р. onkoides. На флете они поселяются преимущественно под колониями кораллов под макрофитами, в расщелинах флета или на стенках прорезывающих его каналов. Максимумы их распространения находятся на подветренной стороне флета и на внешнем склоне на глубинах более 10 м, где воздействие волнения и прямого света ослаблено (Littier, 1973; Doty, 1974).
Аналогичная картина распределения кораллин бьша отмечена на наветренном флете атолла Ронгелап (Маршалловы острова). Начиная от края вся поверхность флета, включая зону "водорослевого вала", целиком покрьпа кораллинами, относившимися к роду Porolithon: Р. onkoides, Р. craspedium, Р. gardineri. При этом Р. onkoides занимал более 50% всей площади его наветренного флета (Doty, Morrison, 1954; Newell, 1956; Doty, 1974; Littler, Doty, 1975). У края наветренного флета тихоокеанских атоллов покрытие площади рифа кораллинами достигает 90-95% (Taylor, 1950; Chevallier et al., 1979). В зоне "водорослевого вала" этих рифов доминируют несколько видов рода Porolithon и Neogoniolithon, и прежде всего Р. onkoides и N. frutescens (Setchell, 1926; Chevallier, Denizot, 1979).
На рифах Атлантики (Сан-Круа, Виргинские острова) структуры, сходные с "водорослевыми валами", строят кораллины с масссивными отростками Lithophyllum congestum (Adey, 1975). Этот вид по своим экологическим свойствам сходен с Porolithon onkoides — главным строителем "водорослевых валов" тихоокеанских рифов. Он поселяется у кромки наветренного рифа, где формирует прочные известковые бугры ("бойлеры"), осушаемые во время отлива. На "водорослевых валах" многих коралловых рифов Индийского океана аналогичную функцию выполняет Neogoniolithon frutescens.
Кораллины, и особенно Porolithon, переносят периодическую осушку при низких отливах лучше, чем талломные макрофиты. Это дает возможность кораллинам полностью занять и надстраивать площадь возвышенных участков флета вблизи его наветренного края, создавая постройки гребня и "водорослевого вала" (Littler, 1973, 1976). Участки флета, где происходит интенсивное выедание макрофитов рыбами и ежами, постепенно занимают корковые кораллины, поскольку последние доступны лишь рыбам-попугаям, тогда как талломные макрофиты и перифитон выедаются большинством всеядных рыб-бентофагов и многими ежами (Van den Hoeck, 1969). Это было доказано экспериментом с установкой на флете клеток, защищающих его поверхность от рыб и ежей. На защищенных участках флет быстро обрастал талломными макрофитами и перифитоном, тогда как на открытых доминировали кораллины (Hixon, Borstoff, 1981). Наконец, кораллины лучше, чем кораллы, выдерживают негативное воздействие загрязнения. Поэтому в зонах, испытывающих загрязнение, кораллы на флете вытесняются кораллинами (Littier, Doty, 1975).
Распределение кораллин на внешнем склоне рифа лимитируется освещенностью. Некоторые кораллины, обитающие у нижней границы звфотической зоны, адаптированы к очень слабому свету. В прозрачных водах они уже обнаруживаются до глубины 400м. Световую адаптацию у кораллин связывают с наличием у них пигмента фикозритрина, полагая, что он способствует поглощению ими коротковолновой радиации в синей области спектра (Littler, 1972). На большинстве рифов они встречаются на их внешнем склоне до глубин 70—100 м. Однако оптимальные световые условия даже для их видов, адаптированных к слабому свету, находятся на глубинах 20—30 м.
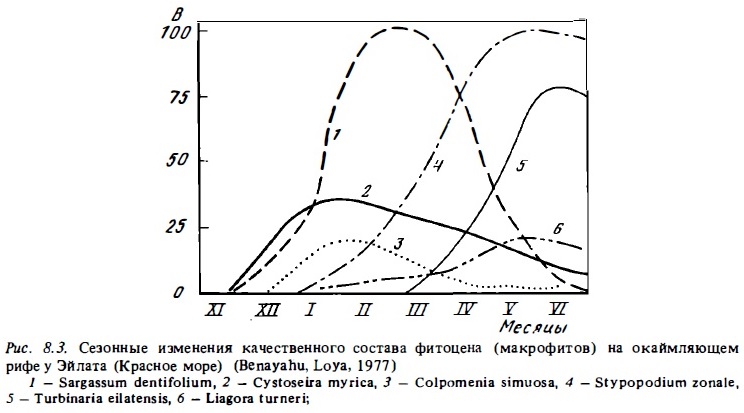
Важная особенность экологии растительных ассоциаций рифа состоит в наличии сук-цессионных изменений их видового состава в донных биотопах (Borowitzka et аl., 1977; Benayahu, Loya, 1977а). Эти сукцессии особенно ярко выражены на высокоширотных рифах, где имеют место сезонные изменения температуры воды. Например, на рифах Эйлата в северной части Красного моря доминирующая в зимнее время ассоциация "Sargassum, Cystoseira, Colpomenia" сменяется летом ассоциацией более теплолюбивых макрофитов — "Stypopodium, Turbinaria, Liagora" (рис. 8.3). При заселении новых биотопов, например искусственна рифов или поверхностей флета, где растительность была уничтожена тайфуном, также наблюдается сукцессионная смена состава поселяющихся на них растительных ассоциаций. В этих случаях вначале поверхность обычно заселяют быстрорастущие нитчатки или зеленые водоросли, такие, как Calotrix, Cladofora, Schaceiaria, Microcoleus, UJva. Эти ассоциации быстро выедаются рыбами, и на их месте возникают ассоциации водорослей, которые имеют более прочные медленно растущие талломы, такие, как кораллины Dictyota, Laurencia, Turbinaria (Tsuda, Kami, 1973; Dollar, 1982) . При занятии кораллами поверхностей флета, где талломные макрофиты интенсивно выедаются рыбами, вначале поселяются быстрорастущие их формы типа Jama, Neogoniolithon. Однако, имея недостаточно прочные талломы, они также довольно быстро выедаются рыбами и ежами и вытесняются кораллинами с более прочными колониями типа Porolithon (Van den Hoeck et аl. 1975).
В составе фитоцена донных биотопов рифа большое значение играют зпифитные водоросли, поселяющиеся на талломах макрофитов и на листьях морских трав. Среди эпифитов есть представители всех типов морских водорослей: диатомовые, синезеленые, бурые и красные. В составе диатомей-эпифитов находятся представители специализированных их групп, обитающих в составе перифитона, на живых талломах водорослей и на кораллах: Auricula, Licomorpha, Rhoralodia, Synedrosphenia.Becьмa распространенными эпифитами, особенно на морских травах, являются нитчатые синезеленые водоросли Oscillatoria, Calotrix, Dichotrix, Plectonema. Среди талломных макрофитов в составе эпифитов преобладают нитчатые формы или вид с мелкими нитевидными талломами размером 1-1,5 см. Из числазеленыхводорослей в составеассоциаций эпифитов встречаются Cladophora, Enteromorpha, Chlorodesmis, Ulvella, Derbesia и из числа бурых водорослей — нитчатые формы Ectocarpus, Sphacelaria, Myrionema, Giffordia. Доминирующую же роль в ассоциации эпифитов чаще всего играют мелкие формы красных водорослей, включая некоторые кораллины. В их числе главными являются родаг Ceramium, Polysiphonia, Hypnea, Chondria, Gelidiella, Griffithsia, Jania, Amphiroa (Fortes 1981; Norris, Bucher, 1982). При этом ассоциации эпифитов отличаются болыпим разнообразием. Так, только на двух видах саргассов, обитающих на рифах у Филиппин, бьшо найдено более 70 видов эпифитов, относящихся ко всем 5 типам водорослей (Chan, 1981).
Морские травы, обитающие на коралловых рифах, предпочитают мелководные ровные прибрежные участки лагун с глубинами в ниэкую воду 0,2-1 м, занятые тонкими коралловыми песками или илистыми осадками. На таких мелководьях они образуют густые заросли типа лугов ("sea grass beds", Zieman, 1975; Tsuda, 1979). Эти заросли состоят обычно из одного или двух видов. В последнем случае один из них доминирует (Zieman et al., 1979). На более глубоких участках дна лагун, а также на песчаных участках флета морские травы образуют смешанные популяции, которые включают макрофиты. На листьях морской травы, особенно на старых и отмирающих в массе, поселяются эпифиты и фораминиферы (Penhale, Smith, 1977; Zieman et аl., 1979).
Для описанных выше лугов морских трав на мелководьях лагун весьма характерным является наличие на них прогалин ("halos") — участков дна, свободных от трав. Эти участки образуются обычно вокруг крупных кораллов, патч-рифов или вдоль кромки рифа, если он граничит с зарослями травы (Randall, 1965). Образование таких зон является результатом выедания травы рыбами и ежами, которые имеют убежища в потч-рифах или под кораллами. Днем они находятся в убежищах, а ночью выходят на "пастбища" и выедают растительность прежде всего с территории, которая находится поближе к их убежищу (Ogden et аl., 1973).