
Большая часть сравнительно крупных рифовых биогенных построек строилась археоциатидами (Вологдин, 1931; Журавлева, Мягкова, 1979; Finks, Hill, 1967; Hill, l972). Их описывают как животных, близких к известковым губкам (Журавлева, 1960), хотя, возможно, некоторые из них представляют собой остатки колоний древних известковых водорослей типа кораллин. Скорее же всего археоциатидные постройки создавались в разные периоды разными организмами со сходной формой скелета: в раннем палеозое — животными типа губок, а позже — известковыми водорослями. Археоциатиды образовывали прочные колонии высотой 10-30 см разной формы: в виде воронок, колпаков, столбиков, чем-то похожих на колонии известковых губок или древних кораллов ругоз. Заполняющий карбонатный материал этих построек формировали в основном строматолиты синезеленых водорослей, а также скелетные остатки губок, моллюсков, трилобитов, иглокожих, мшанок. Расцвет археоциатидных рифоподобных построек приходится на первую половину кембрия (540 млн лет назад). Они имели размеры до 30 м в диаметре (Stubblefield, 1960). Эти постойки напоминали современные патч-рифы. Гряды таких построек формировали на обширных мелководьях теплых морей целые системы типа атоллов или барьерных рифов толщиною до 10м (Яковлев, Рябинин, 1915; Журавлева, 1960; Краснов, 1968; Goldring, 1938; Balzam, 1971; Copper, 1974).
Другой тип рифовых построек раннего и среднего кембрия создавали древнейшие полихеты Scolithos, сходные с современными полихетами сабелляридами и жившими в известняковых трубках, а также губки, сходные с современными губками Demospongia. Кстати, такие рифоподобные постройки эти трубчатые полихеты строят и в наше время (Kirtly, Tanner, 1968). Их размеры могут достигать сотен метров. Автор видел такой полихетный "риф" на побережье Адриатического моря у г. Комаккио близ Венеции. Наконец, в раннем палеозое были распространены рифоподобные постройки, слагавшиеся из биогерм, скелетную основу которых создавали мшанки Trepostoma и Cystoporata, имевшие известковый скелет (Cuffey, 1977а).
Описанные типы древнейших биостромов и рифоподобных построек существовали до раннего ордовика - 470 млн лет назад (Stubblefield, 1960). В раннем ордовике сравнительно небольшие постройки типа патч-рифов формировали известковые губки Archaeoscyphia (сходные с археоциатидами) и Stromatoporaidea, а также зеленые известковые водоросли Celathum (Copper, 1974). Для среднего ордовика характерны' рифоподобные постройки мшанок, а для позднего ордовика — постройки процветавших в то время Stromatoporoidea, животных с прочным известковым скелетом (Lecoinpte, 1956). Они, по-видимому, относились к известковым губкам и были близки к ныне живущим Sclerospongia (Finks, 1970; Hartman, Coreau, 1970; Hartman, 1977). В ордовике появились также первые древнейшие кораллы с прочным скелетом — сначала табуляты, а потом и ругозы (Сошкина, 1947; Соколов, 1955; Ивановский, 1975; Тесаков, 1978). Табуляты характеризовались колониальным ростом и отсутствием выраженных септ (Hill, Stumn, 1956). Эти кораллы вначале имели небольшие размеры колоний — до 30 см высотой. Вместе со строматопоридными губками они формировали арматурную основу рифовых построек. Она заполнялась известковым материалом, который 12 продуцировали синезеленые и зеленые водоросли, а также разнообразные беспозвоночные: мшанки, губки, иглокожие (Manten, 1971; Королюк, Михайлова, 1986). Строматопоридно-табулятные постройки были очень широко распространены в силуре и особенно в девоне (430—340 млн лет назад), когда они достигали внушительных размеров. Их толщина достигала 50—100 м, а диаметр 1—2 км (Lecompte, 1956; Nalivkin, 1967; Brezhnev et al., 1967). В СССР они встречаются в отложениях Урала, Тянь-Шаня (Фане-розойские рифы... , 1986).
В конце силура и в девоне наряду с продолжавшимся развитием строматопорид и табулят появились кораллы ругозы. Они имеют прочные колонии, способные противостоять волнобою (Соколов, 1955). В этот период начался настоящий расцвет процессов рифообразования, в результате чего образовались рифовые постройки площадью в сотни квадратных километров. В зонах тектонического опускания постоянный рост коралловых рифов такого типа в девоне привел к образованию толщ рифовых известняков мощностью до 1600 м, например, на Урале (Наливкин, 1967). Крупные рифовые комплексы сформировались в этот период в Европе (Krebs, 1974), в Австралии (Playford, 1967) и Канаде (Jamisond, 1971). В конце среднего девона и в верхнем девоне в виду наступившего похолодания и понижения уровня океана произошло катастрофическое вымирание кораллов и другой тепловодной фауны (Copper, 1974). В перми и триасе рост рифов возобновился главным образом за счет развития известковых губок Sphinctoidae и Pharetronia (Wilkinson, 1983). Важным, а иногда и преобладающим компонентом многих рифов этого периода были филлоидные красные и зеленые водоросли, близкие к коралиинам и халимеда. Они продуцировали в большом количестве карбонатный материал в виде чешуек и гранул (Wray, 1971). Многие крупные рифоподобные постройки позднего девона — ранней перми толщиною до 50 м состояли почти целиком из этого водорослевого известника (Peterson, Ohlen, 1963; Hecke!, 1974). В процессах рифообразования, протекавших в триасе, основную роль играли наряду с кораллами гидроиды, мшанки, плеченогие моллюски и красные известковые водоросли — кораллины.
В середине триаса появились первые герматипные склерактинии, содержавшие симбиотические зооксантеллы, ареал обитания которых был ограничен теплыми водами. В зонах умеренных и холодных вод в то же время появились агерматипные семейства склерактиний. В числе склерактиниевых кораллов, появившихся в эту эпоху, были как одиночные, так и колониальные формы (Соколов, 1955, 1982; Краснов, 1976; Wells, 1956; Фанерозойские рифы, 1986). Древнейшими среди них были Thamnasteriidae, Astrocoeniidae, Montlivaltiidae, Conophylliidae, Actinacididae - Согласно Е.В. Краснову (1976), разные семейства ископаемых склерактиниевых кораллов произошли от разных групп ругоз. Однако, по-видимому, более обоснованны представления о том, что древние герматипные и агерматипные кораллы, равно как табуляты и ругозы, являются самостоятельными ветвями развития древних полипоидных актиний (Wells, 1956).
Склерактинии впервые получили широкое распространение в составе прибрежной фауны бассейна Тетиса, после чего они довольно быстро становятся важным компонентом фауны всего тропического шельфа (Potts, 1983). В середине юрского периода происходила интенсивная эволюция как герматипных, так и агерматипных склерактиний, в результате чего появилось много новых их семейств. К концу юрского периода много семейств древнейших склерактиний среднего триаса вымирают, дав начало новым, более современным и совершенным семействам склерактиний. У большинства вымерших склерактиний триаса и юры септы кораллитов были не пластинчатыми, как у современных кораллов, а бугорчатыми или шиповатыми (Краснов, 1978). В конце третичного периода произошла дифференциация фауны склерактиний на видовом уровне на индотихоокеанскую и атлантическую (Frost, 1977).
В середине юры в тропической зоне процветали герматипные кораллы, а в умеренных и холодных водах — агерматипные их виды. Эти кораллы в верхней юре создавали мощные рифовые постройки на обширных пространствах тропического шельфа, равно как и на шельфе умеренных бассейнов. В юрский период в некоторых регионах крупные рифовые постройки (до 50 м толщиной) были созданы полихетами сабелляридами и плеченогими моллюсками. Следующий период интенсивного видообразования кораллов и роста рифов имел место в карбоне (Обручев, 1926; Beauvais, Beauvais, 1974). В верхнем карбоне появляются представители семейства сов ременных герматипных склерактиний (Pocilloporidae, Faviidae, Poritidae) и вымирают такие семейства массовых ископаемых кораллов, как Microsolenidae и Amphiasteridae. Значительную роль в формировании рифов начиная с юры приобретают гидроиды: гидроактиноиды (Ellipsactinia), гидрокораллы миллепориды и стиластериды. Расцвет развития последних приходился на поздний карбон (Boshma, 1956).
В палеоцене и эоцене (третичный период) рост коралловых рифов опять замедлился. Продолжалось вымирание ряда семейств древних склерактиниевых кораллов. Вместо них появились семейства современных кораллов Mussidae, Acroporidae, Agarici-dae, Siderasteridae, Fungiidae, Dendrophylliidae. В олигоцене и миоцене интенсивный рост кораллов вновь возобновился с участием представителей семейств склерактиний, родственных современным. Коралловые рифы, сформировавшее в этот период, приобретают вид и строение, близкое к современным рифовым постройкам. Их толщина в некоторых районах достигает 600 м. Наряду с кораллами большую роль в их формировании играли кораллины и фораминиферы. Многие крупные рифовые структуры нииоцена в Северной Америке и в Европе состоят практически из материалов этих компонентов (Howe, 1934). Интенсивный рост толщ рифовых построек на морских мелководьях тропической зоны продолжался и в плейстоцене с некоторыми перерывами в периоды резких понижений уровня океана, связанных главным образом с периодическими оледенениями.
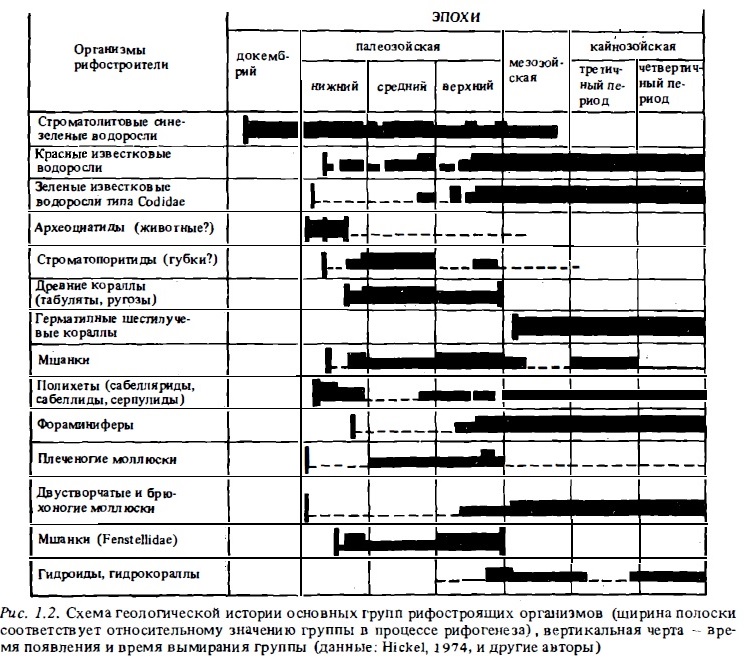
Сведения о геологической истории основных групп организмов-рифостроителей обобщены на схеме (рис. 1.2).