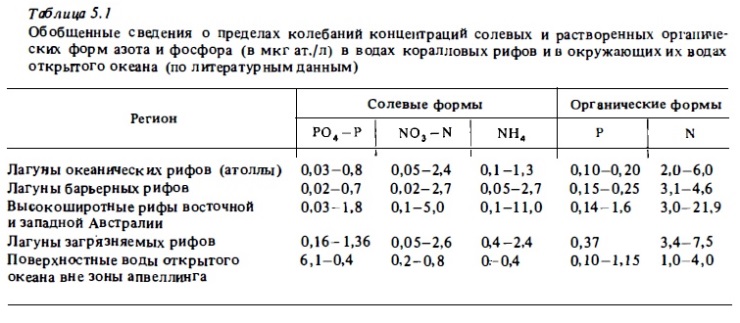
Анализируя данные табл.5.1, нетрудно видеть, что пределы колебаний концентрации солевых биогенов мало меняются на разный рифах, расположенных в разных по уровню продуктивности водах и даже на загрязненных рифах (Smith, 1985). Эти данные показывают, что концентрация солевых биогенов является в значительной степени равновесной величиной, которая отражает лишь направленность процесса круговорота биогенов в момент проведения измерений, но мало говорит об их запасе или об обеспеченности биогенами автотрофных сообществ рифа.
Концентрация минерального фосфата в водах лагун атоллов и барьерных рифов (см. табл.5.1) бывает близкой или несколько превышает содержание РО4—Р в окружающих поверхностных водах океана. В среднем она составляет 0,1—0,4 мкг ат./л, что ненамного выше уровня лимитирования роста фитопланктона (около 0,2 мкг ат./л). Поэтому фосфат, как правило, является основным лимитирующим биогеном в планктонный сообществах рифа. Что касается донных автотрофов, то они, по-видимому, вообще не испытывают биогенного лимитирования, поскольку они получают необходимые биогены преимущественно из их депо и зон регенерации, которые находятся в донный биотопах рифа (Entsch et al., 1983; Smith, 1984; Wiehe, 1985).
Общее содержание солевого азота в форме нитратов, нитритов и аммония в водах, омывающих рифы, и в водах лагун атоллов и барьерных: рифов бывает, как правило, вьине, чем в окружающих поверхностный водах океана (Webb et al., 1975; Andrews, Muller, 1983; Szamant, Froelich, 1983). В среднем оно составляет 0,6—1,5 мкг ат./л, что выше уровня лимитирования роста океанского фитопланктона, который близок к 0,5 мкг ат./л. Более половины имеющегося в водах рифа запаса солевого азота представлено аммонием, тогда как в водах открытого океана аммоний обычно присутствует в очень малый количествах:, а часто не обнаруживается вовсе (Hatcher, Hatcher, 1981). Эти данные говорят в пользу того, что избыток аммония, присутствующий в водах рифа, образуется самими его сообществами за счет процессов азот-фиксации, бактериальной аммонификации и метаболизма фауны. Наибольшие концентрации аммония — до 9-11 мкг ат./л — были зарегистрированы в водах высокоширотный рифов Аброльхос, где имеет место мощное развитие макрофитов и образуется много растительного детрита (Johannes et al., 1983, 1983а), а также в водах лагуны и флета рифа Уантри на юге Б. барьерного рифа Австралии, имеющего мелководную лагуну с интенсивным развитием в ней и на флете макрофитов и перифитона (Hatcher, Hatcher, 1981). В мелководный лужах, остающихся на флете в период отлива и заросших макрофитами и перифитоном, концентрация аммонийного азота достигала 16 мкг ат./л при содержании нитратного азота всего 0,8 мкг ат./л. Эти данные показывают, что донное сообщество рифа производит за счет азотфиксации и выделяет в воду в большом количестве аммонийный азот, который нитрифицирующая микрофлора рифа, ассоциированная в основном с перифитоном и верхним слоем донных осадков, окисляет до нитратов (Webb, Wiebe, 1975; Webb et al., 1975; Atkinson, 1981, 1983).
Абсолютные концентрации солевые биогенов, как правило, мало меняются в разных участках рифа (Odum, Odum, 1955; Pilson, Betzer, 1973). Значительное увеличение их концентрации (до 1 мкг ат./л и более РО4—Р) отмечалось в лагунах барьерных рифов в сезон дождей под влиянием речного стока (Ricard, 1981) и в лагунах атоллов, расположенные в зоне подъема глубинных вод у экватора (до 3 мкг ат./л солевого азота; Kimmerer, Walsh, 1981). Существенное значение в зкосистеме рифа имеют растворенные органические формы биогенов, которые обычно редко учитываются при анализе биогенного режима морских экосистем. Запас растворенного органического фосфата в водах рифов соизмерим с запасом минерального фосфата и составляет 0,1-0,2 мкг ат./л, т.е. примерно столько же, сколько в водах открытого океана (см. табл. 5.1). Судя по имеющимся данным, запас растворенных форм азота в водах рифов в среднем в 1,5-2 раза выше, чем в водах открытого океана. Органические формы биогенов находятся в морской воде преимущественно в виде трудноминералиэуемых соединений. Фосфор присутствует главным образом в составе нуклеиновые кислот и фосфолипидов, а азот - в составе гуминового ядра водного гумуса и ассоциированные с гумусом белков и аминокислот. В водах открытого океана использование органических форм биогенов планктонными организмами затруднено тем, что для их утилизации необходима тесная ассоциация микроорганизмов с макромолекулами, которая облегчает действие их экзоферментов. Формирование таких ассоциаций возможно на поверхностях раздела фаз и на частицах детрита. Однако время жизни детритных частиц на планктонном сообществе ограничено в виду высокой фильтрационной активности населяющего его зоопланктона. На рифе активные микробные сообщества перифитона покрывают обширные поверхности пористого известкового субстрата и кораллового песка. Энергетической основной существования этих перифитонных сообществ служит растворенное органическое вещество вод океана, проходящих нац рифом. Используя это органическое вещество, перифитон получает содержащиеся в нем биогены для собственного биосинтеза, включая их таким путем в общий пул биогенов экосистемы рифа (Сорокин, 1980).
Запас биогенных элементов во взвешенном органическом веществе можно оценить в 0,03-0,06 мкг ат.Р и 0,5-1,8 мкг ат.N (Crossland, Barnes, 1983). Несмотря на относительно небольшую величину этого запаса, его роль в экосистеме рифа можно оценить как первостепенную в связи с тем, что на рифе существуют мощные ассоциации фильтрующих донных животных. Они отфильтровывают органическую взвесь из проходящих над рифом вод океана, включая содержащиеся в ней азот и фосфор, в биогенный пул экосистемы. Часть их они выделяют в виде фосфата, аммония и мочевины, а часть используют для собственного биосинтеза.