
Эррантные полихеты представлены на рифах следующими основными их семействами: Nereidae, Eunicidae, Syllidae, Polynoidae, Amphinomidae. Реже встречаются представители сем. Cirratulidae, Aphrodi tidae, Glyceridae, Phillodocidae.
Нереиды — относительно крупные хищные полихеты. Они имеют умощенное тело с хорошо развитыми параподиями. Захват пищи происходит у них с помощью выдвигающейся глотки и массивных челюстей. Они населяют биотопы с рыхлым грунтом или обломочным материалом, в котором они строят свои норки. Стенки их норок армированы слизью. Червь питается, вылезая почти целиком из норки и поедая все, что он находит вокруг нее: микрофитобентос, детрит, макрофиты, а также и животную пищу. Собрав все съедобное вокруг себя, червь покидает норку и, найдя подходящее место, быстро строит новую. Среди нереид отмечены отдельные случаи трофической специализации. Известны нереиды-фильтраторы (Nereis diversicolor), которые строят, подобно хирономидам, ловчую слизистую сеть в своей норке. Есть среди них специализированные хищники (N.grubel) и фитофаги (N.procera). Однако основная часть нереид представлена всеядными их видами. Некоторые нереиды в период размножения меняют форму тела. Их параподии удлиняются, а на голове появляются крупные глаза, которые занимают почти всю ее поверхность. Таких червей в брачном наряде называют гетероне-рейсами. После метаморфоза они поднимаются в толщу воды, образуя плотные рои, в которых самцы и самки после совершения брачного "танца" выметывают яйца и сперму в воду, где происходит оплодотворение. Большая часть отнерестившихся червей поедается рыбами. Массовыми представителями этого семейства являются: Nereis caudata, N.falcariа, N.jacksoni, N.trifasciata, Platynereis isocita, Perinereis calodonta, P.anomala, Ceratonereis mirabilis.
Эуницицы также представлены достаточно крупными червями, длинными, с круглым телом до 1-2 см в диаметре, с короткими параподиями и бахромой жабр на дорзальной стороне. Большинство эуницид рифа живет в биотопах скального флета в составе криптофауны в отмерших кораллах, под камнями, в пустотах скального флета. Многие эуниниды имеют очень колоритную окраску. У зуницид хорошо развит ротовой аппарат. Они располагают мощной выдвигающейся глоткой. Их челюсти вооружены двойными мандибулами и усажены рядом максилл. Некоторые их виды способны сверлить ходы даже в твердом известняке и в живых кораллах (Lysidice, Palola, Eunice schemacephala). Молодые особи большей части видов эуницид свободно передвигаются по субстрату. Взрослые же особи ведут оседлый образ жизни. Одни виды строят трубки, другие живут в брошенный норках, просверленных сипункулидами, третьи делают собственные норки в рыхлом и даже в скальном грунте (Dry, 1967). Среди эуницид много комменсалов, сожительствуюдых с губками, моллюсками, голотуриями, гидроидами. Есть среди них и паразиты (Arabella). Некоторые их виды (Eunice viridis) образуют в период размножения брачные рои, аналогичные описанным выше роям нереид. В составе эунинид имеются специализированные хищники, которые питаются другими червями, остракодами, мелкими моллюсками (E.aphroditois), а также фитофаги, которые питаются кораллинами и перифитоном (Е. viridis). Большинство же эунинид всеядны с преобпаданием в их рационе животной пищи (Day, 1967; Faucald, Jumars, 1979). Массовыми представителями эунинид на рифах являются: Eunice atra, E.cincta, E.australis, E.antennata, E.caribbea, Palola siciliensis, Lysidice collaris, L.ninetta, Nematonereis unicornis, Drilonereis falcata, Lumhrinereis etraura, Marphysa regulis, M.corallina, Arabella mutans, A.iricolor.
Семейство силлид объединяет мелких полихет размером от 2 мм до 2 см. Они имеют уплощенное тело с тонкими волосковидными параподнями. Это одна из важнейших групп мейо- и микробентоса, которая части доминирует в составе сообществ скального флета, обломочного материала и рыхлых осадков рифа (Kohn, Lloyd, 1973; Hutchings, 1974). Их мощная выдвигающаяся глотка снабжена засасывающим органом — провентрикулом, с помощью которого они высасывают полипы гидроидов. Челюстной аппарат большинства силлид образован одним крупным скребущим зубом и серией мелких зубчиков. Силлиды в большинстве своем не строят временных трубок. Лишь некоторые их виды — комменсалы гидроидов строят такие трубки из слизи, прикрепляя их к стволам своих "хозяев". Большинство силлид — специализированные хищники, пищей которых являются гидроиды, мшанки, асцидии, губки, зоантарии. Однако среди них есть и всеядные формы (род Eusyllis). Эусиллисы живут в песчаных грунтах. В состав рациона этих силлид наряду с животной пищей (остракодами, микромоллюсками) входит микрофитобентос. Важная особенность силлид состоит в их способности к вегетативному размножению. Тело червя делится при этом на куски, на коипах которых регенерируют новые головы и хвосты. Благодаря этому свойству силлиды быстро осваивают субстрат и дают высокую численность (Hutchings, 1974). Доминирующие виды силлид в биотопах рифов представляют: Syllis cornuta, S. alternata, S. spongicola, S.amica, S.gracilis, S.exilis, Opistosyllis laevis, Tryanosyllis zebra, Exogene longicorni s, E.dispar, Е. verugira, Brandia rhopalophora, Odontosyllis ctenostoma.
Полиноиды — относительно крупные полихеты, тело которых покрыто ярко раскрашенными чешуйками. Они живут под камнями и входят в состав криптофауны. Полиноиды — хищники, питающиеся преимущественно другими червями и кладками моллюсков. Яйца оплодотворяются самцом в момент их откладки самой под чешуйки, где и происходит их созревание. Массовые их вид: Harmothoe indica, H.dictyophora, Lepidonotus carinulatus, L.glaucus, L.jukesi, L.jacksoni, Dendropoma sp.
Амфиномиды или "огненные черви" — толстые, крупные, малоподвижные полихеты, живущие под камнями на флете. Их тело покрыто, как шубой, тончайшими ядовитыми ломкими иглами, которые служат им надежной защитой. Амфиномиды-хищники. Их массовые виды: Euri thoe complanata, E.chilensis, Amphinome flava, A.rostrata, Notopygos megalops, Hermodice carunculata. Последний вид известен как кораллофаг, поедающий и сверлящий живые кораллы иослабляющий их устойчивость к волнобою (Marsden, 1962).
Седентарные полихеты представлены на рифах основными их семействами: Serpulidae, Sabellidae, Spionidae, Terebellidae, Capitellidae, Chaetopteridae. Большинство из них — тонкие фильтраторы, способные отфильтровывать бактериопланктон. В состав серпулид как подсемейство входят спиробрисы — одна из наиболее распространенных групп рифовых седентарных полихет. Их прочные армированные известью трубки диаметром до 1,5 см встроены в колонии кораллов. Личинки спиробрисов оседают на поврежденные рыбами или на отмершие участки колоний молодых кораллов. Трубка полихеты растет вместе с колонией, слегка обгоняя ее так, что края трубки всегда приподняты над ее поверхностью. Таким путем червь избегает атаки со стороны полипов коралла-хозяина. На головном конце полихеты находится сложный фильтрационный аппарат в виде двух спиралей, имеющих вид опрокинутых конусов. Он образован ротовыми щупальцами, которые покрыты плотным слоем ресничного эпителия. Поверхность щупалец секретирует тончайшую слизистую сеть, которая, взаимодействуя с потоком воды, возникающим под воздействие ресничного эпителия, вылавливает из нее взвешенные частиц: бактерии, детрит, фитопланктон. Фильтрационные конусы спиробрисов имеют разнообразную поражающую воображение окраску. Они часто десятками располагаются на поверхности колоний крупных массивных кораллов, таких, как Porites или Mentastrea, и служат одним из главных украшений кораллового рифа. В числе массовых видов спиробрисов Spirobranchus gigantea, Spirobris nauti-loides. Другие серпулиды строят прочные извилистые известковые трубки диаметром 1-3 мм на вертикальных скальных поверхностях рифа в зонах субпиторали, флета и склонов. Их длина 0,5-3 см. Иногда трубки серпулид покрывают скальные поверхности сплошным слоем. Серпулида представлены на рифах массовыми видами: Hydroides longispinosa, H.exaltatus, Pomatoceros sp., Serpula vermicularis.
Сабеллиды в большинстве своем - крупные полихеты, которые заселяют преимущественно рыхлые осадки. Из песчинок, склееных слизью, они строят довольно прочные трубки, нижние конусы которых уходят в толщу осадка или под камни, если полихета живет на скальном флете. На рыхлых грунтах они часто образуют плотные поселения. Их фильтрационный аппарат состоит из прямых и длинных перистых щупалец. В случае опасности они мгновенно прячут их в трубку. Массовыми их видами на рифах являются.-Megalomma vcsiculatum, Mbinoculatum, Sabella microphthalma, S.melanostigma, Hypsicomus phaetothaenia.
Спиониды строят постоянные или временные трубки или норки в рыхлом грунте. Некоторые вида (Polydora) способны высверлить норки в мертвых кораллах. Их головной конец снабжен тонкими гибкими щупальцами, поверхность которых покрыта ресничками. С их, помощью червь, высунувшись из норки, собирает с окружающего участка дна осевший детрит. Спиониды способны также и к фильтрации. В случае необходимости они покидают свои трубки и строят новые. Их массовые виды: Malacoceros indicus, Prionospio cirrifera, Polydora armata, P.antennata, Pseudopolydora reishi, Boccardi а polyd ranchia.
Теребеллиды по образу жизни сходны со спионидами. Они строят свои домики-трубки в мягких грунтах под камнями или в колониях мертвых кораллов. Для них характерны очень длинные многочисленные щупальца, снабженные ресничным эннтелием. Располагаясь веером по поверхности дна, черви с помощью ресничек и слизистых тяжей собирают с нее осевший детрит и гонят его в сторону рта, где с помощью специальных выростов-губ пищевые частицы сортируются, а затем заглатываются. Массовые виды теребеллид: Terebella ehrenbervi, Liomia medusa, Streblosoma hesslei, Hanchiella tribullata, Terebellidae strolmi, Eupolimnia nebulosa, Reteterebella queen-slandia.
Капителлиды по форме напоминают дождевых червей. Из слизи и песка они строят себе трубки в рыхлом осадке, с поверхности которого они собирают детрит и микроводоросли. Их массовые вида: Scyphoproctus somalus, Dasybranchus caducus, D.cadutus, Pulliclla armata.
Хетоптериды живут в трубках, погруженных в рыхлый осадок или укрепленных в щелях твердого грунта. Питаются они, прогоняя воду с помощью ресничного аппарата через слизевую сеть, выделяемую параподиями. Образующийся пищевой комок затем заглатывается (Barnes, 1964). На рифах распространены: Chaetopterus variope-datus, Mesochaetopterus minutus, Phyllochaetopterus sp. Остальные семейства полихет представлены массовыми их родами: Cirriformia, Dodecaceria, Cirratulus, Cirriformia (сем. Cirratulidae}, Phyllodoce, Eulalia, Notophyllum, Pterocyrrus (сем. Phyllo-docidae), Armandia, Ophelia (сем. Opheliidae).
Как мы видели выше, среди полихет есть сидячие фильтраторы, сидячие и бродячие детритофаги и сестонофаги, хищники и паразитические виды. Они в большом количестве заселяют норы, щели и пустоты скального флета, обломочный материал и рыхлые осадки. Полихеты являются важным компонентом криптофауны отмерших кораллов. При анализе криптофауны только одной большой колонии отмершего коралла на Большом барьерном рифе было найдено более 100 видов полихет (Grassle, 1973). Многие полихеты живут в сожительстве с губками, кораллами и гидроидами, являясь их комменсалами или симбионтами. Они играют существенную роль в плаиктоне рифа (Sale, 1976). Многие мелкие полихеты периодически выходят в толщу вод рифа. В большом числе донные полихеты появляются в толще вода в период нереста. Наконец, важным элементом меропланктона рифа являются их личинки трохофоры. Наряду с этим среди полихет есть и чисто планктонные их формы, такие, как томоптериды.
Учет фауны полихет в биотопах скального флета, где они играют доминирующую роль в составе зообентоса, сопряжен с большими трудностями. Исследователю приходится вырубать зубилом блоки скального известняка. Эта бирка затем фиксируют формалином, после чего их дробят и вымывают из них фауну (Kohn, Lloyd, 1973; Hutchings, 1974; Vittor Johnson, 1977) или полностью растворяют куски известняка в соляной кислоте (Hutchings, Weate, 1978). При таком способе отбора проб скального материала часть подвижной фауны может уйти. В связи с этим при оценке имеющихся данных об общем числе видов и о численности и биомассе полихет следует учитывать, что они могут быть существенно заниженными (Fauchald, 1974). В последние годы бьт найден метод, обеспечивающий более полный учет фауны полихет в скальном субстрате. Он состоит в установке на дне блоков пористого кораллового известняка объемом около 1 дм3 на срок 2-3 года (Hutchings, 1981, 1983, 1985). За этот период блок заселяют полихеты, обитатели окрестного биотопа. В таких блоках удается обнаружить больше видов полихет, чем в пробах, вырубленных рядом из скального флета. Так, в блоках, установленный на рифе у о-ва Лизард на срок 2 года, бьто обнаружено 144 вида полихет, что намного выше, чем можно бьто обнаружить в пробах. В блоках преобладали полихеты Polydora, Syllis, Hydroides, Ceratonereis, Nematonereis. Рекрутирование полихет идет в основном путем оседания из водной толщи их личинок и молоди. Важное значение имеет первоначальное поселение на блоках сверлящих сипункулид, высверливающих норки, которые затем заселяют полихеты.
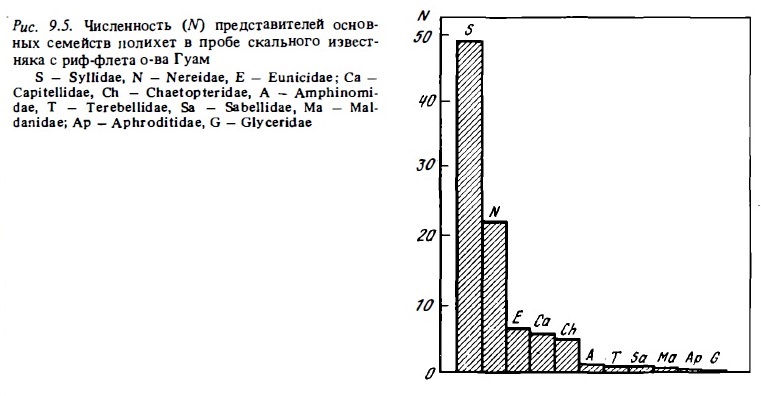
Общее число видов полихет, заселяющих отдельные рифы, достаточно велико и может достигать 100-130 (Западная Атлантика). На багамских рифах обнаружено 46 видов, на рифах Тулеара — 118, на рифах Гуама — 29, на рифах Таиланда и Суматры — 76, на атоллах французской Полинезии 132 вида, в южной части Б .барьерного рифа — около 140 видов (Kohn, Uoyd, 1973; Hutchings, 1974, 1985; Vittor, Johnson, 1977; Peyrot-Clausade, 1977, Devaney, 1987). По общему числу видов в большинстве случаев преобладают представители сем. Syllidae и Eunicidae. По общей численности доминируют эти же семейства, а иногда также и серпулиды. По биомассе обычно доминируют эунициды, нереиды и капителлидкг Доминирующее значение силлид обеспечивается в значительной степени тем, что они способны к вегетативному размножению. Анализ структуры сообществ полихет на ино-тихоокеанских рифах показывает, что в биотопах скального флета, покрытого живыми и отмершими кораллами, макрофитами и кароллинами, равно как и в мелководный донных биотопах, занятых обломочным материалом, как по числу видов (10—20), так и по численности (10—60 тыс/м2),обьто доминируют мелкие силлид размером 2-15 мм. За силлидами следуют эунициды, капителлиты, нереиды, хетоптериды (КоЫл, Lloyd, 1973; Hutchings, 1974, 1985; Peyrot-Clausade, 1977; Бужинская и др., 1980) (рис. 9.5). На рифах Западной Атлантики в этих биотопах преобладают эуницид и серпулида (Vitton, Johnson, 1977). Сидячие полихеты ( в основном серпулиды) часто доминируют в сообществах каменистой литорали. Индекс видового разнообразия Шеннона Н' в сообществах полихет скального флета достаточно высок: 1,5-2,8.
В мелководных биотопах рифа, занятых рыхлыми осадками, заросшими травой или макрофитами, а также на дне лагун фауна полихет несколько беднее, чем на твердак грунтах рифа, хотя ее значение в формировании бентоса рыхлых осадков остается весьма высоким. Средняя численность полихет в этих биотопах составляет 5-20 мыс/м2 при биомассе 10—100 г/м2 (Thomassin et al., 1976, 1982) . В составе доминирующих здесь таксонов эррантных полихет эунициды (Marphysa), капитеплиды (Dasybranchus), полиноиды (Lepidoiiotus), нереиды (Nereis, Perinereis), глицериды (Glycera). Большое значение в этих биотопах имеют сецентарные полихеты хетоптерусы (Chaetopterus, Me-sochaetopterus), теребеллид (Terebellidaes), спионид (Prionospio). Структура сообществ полихет зависит от условий седиментации и от обилия детрита и илшгктона в толще воды. Так, на прибрежных рифах у Флориды, обогащенных детритом и планктонодоминируют фильтраторыысерпулиши, а на рифах, где планктон беднее, доминируют сестонофаги-эунициды. В целом же в составе сообществ рифов в среднем около 30% общего числа видов полихет представлено фильтраторами, около 40% — детритофа-гами и около 30% — хтцниками (Vittor, Johnson, 1977).
Общая численность полихет в мелководных донных биотопах, по оценкам разных авторов, варьирует в пределах 10-130 тыс./м2 при биомассе от 20 до 200 г/м2 (Hutchings, 1974; Kohn, White, 1977; Reichelt, 1979). Последняя величина следует из данных об биомассе полихет (преимущественно эуницид) на рифах у Тулеара, равной 0,9 r сухого веса на 1 дм3 скального известняка (Peyrot-Clausade, 1977). Тогда при условии, что сухой вес полихет равен 20% от сырого в слое 0—5 см, их биомасса и будет выражаться приведенной выше величиной - более 200 /м2. При этом, как отмечалось выше, 50—70%, а иногда и 90% общей их численности составляют силлиши. Однако их доля в биомассе обычно меньше, чем у остальных полихет, ввиду их малых размеров (2-10 мм) и малого веса (0,2—2 мг/экз). По биомассе же доминируют чаще всего эуницидн1, капителли-дысерпулиды и реже нереиды.
Полихеты по своему значеиню в трофодинамике рифа занимают одно из ведущих мест в сообществах рифа. Они используют практически все доступные источники пиши и создают при этом большую биомассу, обладающую высокой кормовой ценностью. Они служат важнейшим источником питания большинства рыб-бентофагов, многих рыб-планктонофагов и хтцных донных беспозвоночных, особенно хищных моллюсков (Kohn, 1968; Bakus, 1969; Vittor, Johnson, 1977). Имея малые размеры и высокую скорость роста, обладая частично способностью к вегетативному размножению, полихеты создают существенную, а во многих случаях преобладающую долю продукции всей донной фауны (Hutchings, 1974, 1981). Обладая способностью сверлить норы в скальном известняке, многие полихеты участвуют в процессе биоэрозии рифа (Hutchings, 1974; Warme, 1977).
Наряду с полихетами существенную роль в донных биотопах рифа играют и другие группы червей, среди которых выделяются сипункулиднт и нематоды. Эхиуриды, оли-гохеты и турбеллярии также являются обычными компонентами донной фауны в некоторых биотопах рифа, однако их численность, как правило, сравнительно невелика.
Сииункулиши и эхиуриды. Сипункулид и эхиурид выделяют в настоящее время в отдельные типы беспозвоночных. Сипункулиды имеют короткое мешковидное или цилиндрическое несегментированное тело размером от 2-3 мм до 10 см, которое оканчивается втягивающимся хоботком. Они встречаются во всех типах грунтов рифа, высверливая или выкапывая себе убежища в виде норки. Многие их виши способны сверлить коралловый известняк (Warme, 1977). Рот сипункулид находится на конце растяжимого и подвижного хоботка, длина которого часто соизмерима с длиной тела. Ротовое отверстие окружено венчиком щупалец. Конец хоботка часто вооружен роговыми зубчиками. Численность сипункулид может достигать значительных величин. Она максимальна в зоне гребня флета - до 1-2 тыс./м2. В рыхлых грунтах их численность невелика: 20-50 экз/м2 (Rice, Macintyre, 1982). Биомассу их в зонах скального флета можно оценить в 10-30 г/м2. Сипункулиды служат источником питания рыб-бентофа-гов и особенно хищньк моллюсков, таких, как конусы и митры. На отдельных рифах число их видов варьирует от 5 до 30. Всего же на рифах обитает до 300 видов сипункулид (Rice, 1976). В биотопах скального флета преобладают сипункулиды, способные сверлить известняк: Paraspidosiphon, Aspidosiphon, Lithacrosiphon. Червь Aspidosi-phon elegante сверлит норки в живых кораллах, а A.jukesi живет в симбиозе с одиночным кораллом Heterocyathus. Сипункулид Siphonosoma vasum, S.no-vaepommeraniae, Phascolosoma perlucens, Golfingia misakiana роют норки в рыхлом грунте.
Эхиуриды по внешним чертам строения напоминают сипункулид. От их мешковидного тела размером 0,5-5 см отходит хоботок, который у них в отличие от сипункулид не втягивается внутрь тела. Рот находится на конне хоботка, вооруженного крючками. Эхиуриды живут в норках, в щелях, под камнями. С помощью своего хоботка они собирают оседающий поблизости детрит. На рифах они представлены родами: Anelas-sorhynchus, Pseudobonellia, Achaetobonella.
Олигохеты. Эта группа кольчатых червей-ашелид представлена мелкими тубифици-дами и в меньшей степени энхитреидами. Олигохеты в сравнительно небольшом количестве входят в состав мейобентоса рыхлых осадков лагуны и особенно литорали барьерньк и окаймлдкщих рифов, куда поступает детрит с суши. Их значение может возрастать в зонах восстановленных осадков и заиления. Олигохеты — одна из наименее изученных групп мейобентоса. Их численность в эонах наибольшего обилия составляет 50-200 экз/дм3 осадка, что на порядок меньше численности нематод (Thomassin et аl., 1982). Размеры их 5-20 мм. Некоторые тубифидида из родов Phallodrilus и Coral-lodrilus не имеют кишечника, но располагают симбиотическими бактериями. Большинство же олигохет питаются микрофлорой и мелким детритом. В составе мейофауны донных осадков рифа наряду с упомянутыми выше родами тубифицид можно встретить роды Limnodriloides, Heterodrilus, Marcusaedrilus (Jamieson, 1977; Mather, Bennett, 1984).
Нематоды. Круглые черви нематода обычны в составе мейобентоса рыхлых грунтов коралловых рифов (Renaud-Mornant et al, 1971, 1971а; Thomassin et al., 1975, 1976; Grelet, 1985). Их биомасса в коралловом песке близка к 1-10 г/м2 при численности 0,2-2 млн/м2. Их размерь! составляют всего 0,2-1 мм и индивидуальный вес 310 мкг. Как видно из приведенньх величин, биомасса нематод в рыхлых осадках рифа весьма значительна и является важным источником питания всевозможной донной фауны и рыб. Сами они в большинстве своем — грунтоеды или детритофаги. Среди них есть и хищники (Salvat, Renant-Morant, 1969).
Немеретины. На рифах в составе инфауны, под камнями и в обломочном материале встречаются представители ленточных червей-немертин из классов Anopla и Enopla. Некоторые их виды достигают 1 м и более в длину (Baseodiscus hempria, B.delineatus, B.guinquelilineatus). Длина других массовых немеретин Gorgonorhynchus repens и Nemerites hermaphroditicus — 5—10 см, а Poseidonemerites bothwelae — 1 см. Мелкие немеретины могут составлять заметную долю мейобентоса, песков. В большинстве своем немеретины — хищники (Denaney, 1987; Gibson, 1982).
Турбеллярии (плоские черви). Свободно живущие плоские черви, обитающие на рифах, относятся преимущественно к отряду Polycladida (роды Pseudoceros, Stylochoplana, Paraplanocera). Они имеют размеры 1—5 см. Многие из них ярко раскрашены. Чаще всего они — комменсалы кораллов, губок или обитатели криптофауны. Их численность на рифах невелика.