
Филогенетически губки занимают промежуточное положение между одноклеточными и многоклеточными животными. Их ткани не имеют дифференцированной эктодермы и эндотермы. Они состоят из специализированных групп клеток, двигательная активность органелл которых в определенной мере синхронизирована, хотя каждая клетка питается самостоятельно. Тело губки состоит из межклеточного структурного вещества (мезглеи), в котором передвигаются амебоидные клетки. Мезоглея покрыта снаружи слоем эпителиальных клеток (пинакодермой). Они же выстилают поверхность каналов, пронизывающих губку. На этих каналах имеются расширения — фильтрационные камеры (рис.9.2). Поверхность камер выстлана слоем специализированных воротничковых клеток, имеющих жгутики. С их помощью в фильтрационных камерах создается постоянный ток воды, из которой хоаноциты с помощью жгутиков, воротничков и слизи извлекают пищевые частицы, осуществляя таким путем фильтрационное питание. С помощью электронной микроскопии было найдено, что воротничок хоаноцита представляет собой подобие сита, состоящего из отдельных нитевидных выростов цитоплазмы, скрепленных поперечными тяжами. Хоаноцит прогоняет с помощью жгута воду через воротничок, как через фильтровальную воронку, которая улавливает взвесь (Rasmont, 1968). Часть уловленных хоаноцитами пищевых частиц фагоцитируют и переваривают они сами, а часть передают другим клеткам. Любопытно, что у хоаноцитов личинок воротнички сплошные, как их себе и представляли раньше. Материал, из которого построена скелетная основа губки - мезоглея, состоит в основном из волокнистого структурного белка - коллагена и из волокон очень прочного рогоподобного вещества - спонгина. Волокна коллагена и спонгина армированы кремневыми или известковыми спикулами. Форма спикул у губок водоспецифична. Они бывают игольчатыми, трех- и четырех лучевыми. Материал спикул известковых губок наряду с кальцитом содержит карбонаты марганца и стронция.
Форма колоний губок очень разнообразна.от инкрустирующих, листовидных и чашевидных форм до трубкообразных и массивных. Их средние размеры составляют у большинства видов 20-50 см. Сверлящие губки имеют меньшие размеры колоний, а губки с трубкообразными колониями могут достигать в высоту 2м. Форма колоний губок имеет адаптивное значение. Инкрустирующие, листовидные и чашевидные колонии с относительно тонкими стенками образуют, как правило, губки, которые живут в симбиозе с водорослями. Тонкие стенки и большая удельная поверхность колонии обеспечивает оптимальные условия освещенности для водорослей-симбионтов, которые служат важным источником питания губки-хозяина. Инкрустирующая форма колоний характерна и для многих видов сверлящих губок. Трубообразная форма колоний обеспечивает лучшие условия фильтрационного питания для губок крупных размеров. В такой копании при ее взаимодействии с течением возникает "тяга", как в печной трубе, что снижает траты энергии губкой на процесс фильтрации (Reiswig, 1974).
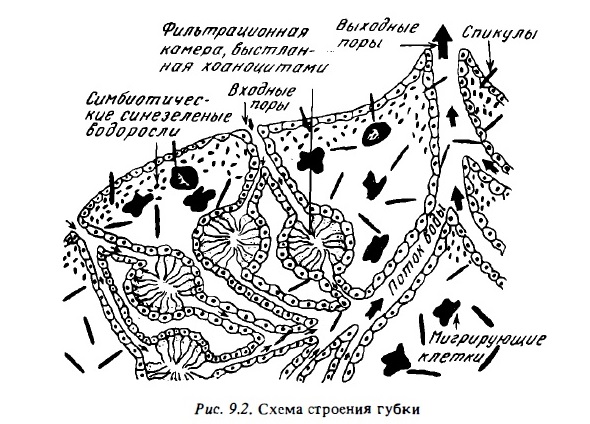
Губки, как и кораллы, размножаются как половым, так и бесполым путем. Среди них есть гермафродиты и двуполые видл1. Оплодотворение, как правило, внутреннее. Во внешнюю среду выделяются оплодотворенные яйца или личинки. Личинки губок очень мелкие. Большинство из них живут в планктоне, хотя есть среди них и ползающие по дну. Вететативное размножение губок происходит почкованием или фрагментированием. Последний способ особенно характерен для сверлящих и инкрустирующих губок, которые разносятся вместе с кусками колоний кораллов, на поверхности которых они растут и затем приживаются на новом месте. Растут осевшие губки довольно быстро. Так, осевшая личинка губки Mycale за год вырастает в губку объемом 50 см³.
Губки (наряду с полихетами, сабеллидами и серпулидами) являются наиболее мощными тонкими фильтраторами в донных сообществах рифа. Синхронно работающие жгутики хоаноцитов создают в каналах, пронизывающих тело губки, постоянный ток воды. При этом крупные частицы взвеси улавливаются на поверхности колонии и в каналах выстилающими их эпителиальными клетками-пинакоцитами. Мелкие частицы, включая бактерии, улавливаются в основном воротничковыми клетками-хоаноцитами в фильтрационных камерах. Бактерии служат главным источником питания губок (Сорокин, 1971; Sorokin, 1973, 1978; Reiswig, 1975). В соответствии с данными последнего автора, скорость фильтрации, определенная путем прямого измерения потоков воды через колонию, близка к 0,5-0,8 л/ч на l см³ объема колонии. Каждае 4-20 с губка профильтровывает объем воды, равный ее собственному объему. Наши измерения показали, что скорость фильтрации у губок близка к 200-400 л вода на 1 г углерода тканей в сутки. При этом губка практически нацело извлекает из вода бактериопланктон и фитоплацктон. Наряду с этим из воды извлекается часть коллоидных частиц и растворенного органического вещества. Энергетические затраты губок на процессе фильтрации, выраженные в единицах поглощенного ими кислорода, составляют в среднем 1-4 мкг О₂ на 1 л воды. При этом в последнем случае при затрате 4 мкг О₂ /л губка (Verongia) пр расчетам не может покрыть трат кислородами на обмен за счет отфильтрованной пищевой взвеси. Дополнительными источниками энергии могут быть у нее фотосинтез ее симбионтов-водорослей и потребление ею растворенной органики (РОВ). Интенсивность дыхания губок выражается величинами 0,02-0,07 см³ О₂/см³ колонии в час, или 0,4-1,5 см³ О₂/г сухого веса колонии в час (Reiswig, 1974). Суммарный рацион сообщества губок в зоне фор-рифа у Ямайки он оценивает в 1,5-4 г органического вещества на 1 м² в сутки. В состав рациона входят все компоненты взвеси (бактерии, фитопланктон, микрозоопланктон, детрит),а также коллоидные частицы и РОВ (Stephens, Shinske, 1961; Reiswig, 1975; Sorokin, 1978; Vacelet, 1979; Wilkinson, Garrone, 1980; Wolkinson, 1978, 1983). За счет фильтрационного питания губки, не содержащие симбионтов (Mycale), покрывают 100% трат на обмен, тогда как ситбионтные (Verongia) всего около 20%. Остальные 80% они получают за счет фотосинтеза симбионтов и усваивая растворенное органическое вещество (Reiswig, 1974).
Способность к симбиозу у большинства губок, заселяющих верхние и средние горизонты рифов, в значительной мере объясняет их способность успешно конкурировать с другой сидячей фауной. Губки в отличие от конечнополостных имеют в своих клетках в качестве эндосимбионтов цианобактерии (синезеленые водоросли) и гетеротрофные бактерии. Присутствие гетеротрофных бактерий в клетках губок было обнаружено еще в 30-х годах. Позже эти наблюдения подтвердились (Levi, Levi, 1965; Vacelet, 1971). Подробными исследованиями было доказано, что зти бактерии являются не паразитами, а эндосимбионтами губок (Wilkinson, 1978, 1978а, 1983; Wilkinson, Garrone, 1980). Бактерии находятся в вакуолях специальных клеток-бактериоцитов и в скелетной массе губки — мезохиле (рис.9.3). Общее их число достигает 10⁷ клеток/г сырого веса, причем у некоторых губок биомасса бактерии составляет до 30-40% всей массы клеток губки (Vacelet, Donadey, 1977; Wilkinson, 1978, 1983). Оказалось, что одна губка может содержать до 5-7 разных видов бактерий.

Исследования видового состава бактерий, населяющих ткани губок, показали, что часть этих бактерий неотличимы от обычных планктонных бактерий. В то же время некоторые бактерии оказались специфичными и присутствовали во многих видах губок, обитающих в разных регионах (Wilkinson et al., 1981). У губок Terpios в мезоглее обнаружены симбиотические бактерии, имеющие нитчатую форму голубой пигмент. Морфологически эти бактерии сходны с нитчатыми цианобактериями, но лишены хлорофилла (Santavy, 1985). Функция бактерий, как эндосимбионитов, состоит главным образом в усвоении растворенного и взвешенного вещества из потока воды. При этом они наращивают свою биомассу, которая затем переваривается клетками губки. Действительно, в присутствии меченного тритием аланина метка появлялась 194 прежде всего в бактериях, находящихся в клетках губки, что было видно на радиоавтографах срезов. Позже метка появилась и в клетках губки (Wilkinson, Garrone, 1980).
Важная функция бактерий-симбионтов, населяющих мезоглею, состоит в том, что бактерии, потребляя излишки коллагена и взвесь, прочищают каналы, пронизывающие тело губки, и поддерживают тем самым ее нормальную фильтрационную активность. В числе массовых видов губок, имеющих в своих тканях бактерий-эндосимбионтов: Paricharax, Jaspes, Neofibularia, Carternspongia. Симбиотические бактерии присутствуют в яйцах некоторых губок и таким путем передаются их потомству (Levi, Levi, 1976). Во многих губках в качестве зндосимбион-тов одновременно присутствуют бактерии и синезеленые водоросли.
Сине зеленые водоросли известны в качестве эндоосимбионтов только у губок и у некоторых червей-эхиурид (Kawaguti, 1971). Впервые эндосимбионты у губок были найдены Фельдманом в 1933 г. Он обнаружил в клетках губок одноклеточные и многоклеточные синезеленые водоросли. Позже у некоторых губок из рода Cliona в качестве эндосимбионтов были обнаружены зооксантеллы и даже криптомонады (Sara, Liaci, 1964), Синезеленые водоросли-симбионты губок представлены у большинства из них мелкими одноклеточными цианобактерий Aphanocapsa feidmanni (реже — А. raspaigclla) — специализированными симбиотическими видами, близкими к пелагическим цианобактериям Synechococcus. Размер клеток А. feldmanni 2—3 мкм и А. raspaigella 6—12 мкм. У некоторых губок (Tethya, Ulosa) в качестве симбионтов присутствует многоклеточная нитчатая синезеленая водоросль Phormidium spongeliae. Синезеленые водоросли-симбионты присутствуют в среднем у половины всех видов губок, заселяющих освещенные зоны рифов (Vacelct, 1981; Wilkinson, Trott, 1985). Они присутствуют в большом количестве в верхнем слое тканей губки толщиною до 1 см в зависимости от их светопроницаемости. Одноклеточные синезеленые-симбионты находятся преимущественно в вакуолях крупных специализированных клеток-цианоцитов. В одной такой клетке их бывает до нескольких сотен (Wilkinson, 1980). Часть их находится также и в мезоглее. Общая биомасса синезеленых водорослей в составе симбиотических губок может достигать 50% биомассы клеток самих губок. Многоклеточные же синезелсные водоросли находятся преимущественно в мезоглее. Цианоциты регулируют размножение симбионтов и их метаболизм так же, как и клетки кораллов, регулируют обмен и размножение зооксантелл. Они стимулируют транслокацию продуктов фотоситеза водорослей, получая от них питание таким биотрофным путем. Избыток клеток симбионтов цианоциты переваривают, получая от них питание также и некротрофным путем (Sara, 1971; Vacelet, 1971). В качестве транслоцируемых водорослями продуктов фотосинтеза преобладают глицерин и гликоген (Wilkinson, 1980, 1983а) . По оценкам последнего автора, транслокация составляет около 12% всей продукции симбиотических водорослей. Остальная их чистая продукция используется губкой некротрофным путем.
До сих пор не вполне ясно, каким путем происходит "заражение" губок симбиотическими синезелеными водорослями. Судя по тому, что они присутствуют в личинках некоторых губок, "заражение" происходит, по-видимому, уже на стадии яйца. Наличие симбиотических водорослей оказывается полезным для губок в нескольких аспектах. Главный из них, несомненно, состоит в получении дополнительной энергии, поставляемой губке-хозяину симбионтами. За счет их фотосинтеза губки могут при оптимальных условиях освещенности компенсировать более половины своих суточных трат на обмен. Другая важная функция симбионтов губок состоит в том, что они, поглощая свет, снижают ингибирующее действие прямой солнечной радиации на клетки губки-хозяина. Наряду с этим синезеленые водоросли обладают способностью потреблять солевые биогены из вода и поставлять их уже в органической форме клеткам хозяина. К тому же симбиотические водоросли потребляют продукты обмена клеток губок и переводят их в состав собственной биомассы, которая затем снова используется клеткой-хозяином. Наконец, синезеленые водоросли-симбионты фиксируют молекулярный азот, улучшая тем самым азотный баланс губки-хозяина (Wilkinson, 1979). Не случайно поэтому губки, которые имеют растительна симбионты, растут на свету намного быстрее, чем в темноте (Wilkinson, Vacelet, 1979).
Синезеленые-симбионты присутствуют в губках разных таксономических групп. Они были найдены у представителей многих семейств известковых губок и склероспонгий. В сообществах губок, обитающих на рифах, можно выделить три их трофические группировки: группу фототрофов, группу миксотрофов и группу гетеротрофов. Эти группировки отличаются по степени участия автотрофной продукции в компенсации их обмена. Она выражается отношением фотосинтеза и дыхания (Pt/Mt), измеряемым в середине дня при среднем уровне освещенности, типичном для глубин 10-30 м, где обитают губки (400 мк Е/ (м²·с) . У фототрофных губок это отношение выше 1,5, у миксотрофных — < 1,5, а у гетеротрофов близко к нулю. Максимальные значения отношения часовых величин Pt/Mt в середине дня (2,5—4) зарегистрированы у симбиотических губок, веерообразных или чашевидных, колонии которых имеют тонкие просвечивающие стенки. В среднем эти фототрофные губки за счет фотосинтеза своих симбионтов компенсируют летом 60—120% своих трат на обмен и зимой 50—70%. Заросли губок в биотопах флета и склонов рифов центральной части Б. барьерного рифа Австралии плотностью около 300 г/м² создают автотрофную продукцию в размере 0,1—0,2 г С/м² (валовый фотосинтез). Это составляет около 2% от валовой первичной продукции рифа, равной 7,7 г С/(м²·сут) (Wilkinson, 1981, 1983а). Измерения зависимости интенсивности фотосинтеза губок от освещенности показали, что ее оптимум лежит в пределах интенсивности света 170—300 мк Е/м²·с), что близко к 50—150 W/м² (рис. 9.4). Это несколько выше оптимума освещенности, характерного для кораллов. Для фототрофных губок отмечена световая адаптация к пониженной освещенности (Wilkinson, 1981; Wilkinson, Trott, 1985). Содержание хлорофилла в колониях губок варьирует у разных видов от 10 до 100 мкг/г сырого веса. Максимум поглощения хлорофилла губок близок к 664 нм (Wilkinson, 1979). Фототрофные виды губок доминировали в сообществах флета некоторых рифов Б. барьеврного рифа. На флете и в верхней части склона рифа Дэвис они составляли 80% всех видов губок. У 7 из 9 массовых видов отношение Pt/Mt было близко к 3 (Wilkinson, 1983).
