Основой динамики биогенов в любой экосистеме является их биотический круговорот. Процессы динамики биогенов, протекающие в толще воды рифов, включают потребление солевых биогенов: фосфатов, нитратов и аммония микропланктоном (фитопланктоном и бактерио планктоном). При этом растворенные солевые биогены переводятся во взвешенную органическую форму, включаясь в состав их биомассы, а также частично в растворенные органические их соединения, которые экскретируются в воду микропланктоном. Процессы, обеспечивающие баланс солевых биогенов в толще воды, включают частичную дыхательную минерализацию их органических форм самим микропланктоном. Основная же их минерализация, перевод из органических соединений в солевую форму осуществляются за счет деструкционной переработки микропланктона в пищевой цепи в процессе его использования консументами-планктонными и донными фильтраторами рифа. До недавнего времени основные усилия исследователей были направлены на изучение динамики биогенов в донньх биотопах рифа, при этом игнорировалось изучение процессов их круговорота, протекающих в толще воды. Однако даже простой расчет, основанный на величинах суточной деструкции в толще воды рифа (0,15—0,3 мг О2/л), дает величину суточной регенерации фосфора 0,6-2 мкг/л, что при его запасе в воде 5—10 мкг/л обеспечивает время его оборота около 5—8 сут и без учета влияния донных сообществ. Учитывая, что интенсивность обмена минерального фосфата между донными биотопами рифа и водной толщей в целом невелика, эту оценку можно было вполне принять в качестве ориентировочной. Тем не менее непосредственные определения скорости потребления биогенов в водной толще, которые позволили бы составить представления о времени оборота из запаса в толще воды, были очень интересны. для выяснения общей интенсивности круговорота биогенов в толще воды рифов и относительной роли в нем бактериопланктона мы выполнили необходимые исследования с применением меченого фосфата. Исследования 82 проводились на атоллах Херон и Вистари зимой и весной 1986 г. с использованием ранее описанной нами методики (Сорокин, 1985). Некоторые из полученных результатов представлены в табл. 5.2. Скорость потребления минерального фосфата микропланктоном в водах исследованных рифов оказалась очень высокой, несмотря на низкую фоновую концентрацию фосфата в воде. Она составляла в среднем около 1 мкг Р/л в сутки при колебаниях от 0,3 до 5 мкг/л. Зимой (в августе) при низкой температуре воды она в целом была ниже, чем весной (в октябре), когда начинался прогрев воды. Сезонную направленность процессов круговорота биогенов отмечали и другие авторы (Hatcher, 1985). Высокие величины часто отмечались в зоне флета во время отлива, а также в водах лагуны и прилегающих к рифам водах открытого моря, когда в гюследних отмечались высокие величины первичной продукции (см. табл. 5.2). В рифовых же водах над флетом величина Paf была часто достаточно высокой и при слабом развитии фитопланктона. Основная его часть (50-90%) потреблялась бактериопланктоном, составляя 50-90% от общего его потребления. Таким образом, бактериопланктон вод коралловых рифов выступает как один из основных агентов в круговороте биогенов. Его функция состоит в потреблении солевых биогенов из проходящих над рифом вод океана за счет использования энергии обедненной биогенами органики, которая поставляется в толщу воды фотосинтезирующими сообществами рифа. Произведенную таким путем биомассу бактериопланктона рифовых вод потребляют донные и планктонные фильтраторы, поставляя содержащиеся в нем биогены в донные биотопы в виде фекалий и растворенных фракций солевого и органического азотаи фосфора.
В условиях относительной стабильности концентрации РО4—Р в водах рифа и при сравнительно низкой скорости обмена фосфором между водной толщей и донными осадками величина потребления фосфата планктонным сообществом должна быть в значительной мере сбалансирована с процессом его новообразования в ходе деструкции органического вещества в водной толще, поскольку основной его круговорот должен протекать в этих условиях в толще воды. Попытка расчета интенсивности минерализации на основе полученных нами в тот же период данных о суточный величинах деструкции органического вещества (D) известного отношения С:Р, близкого к 40 (по весу), дает вероятную величину скорости минерализации фосфора в толще воды 1-3 мкг Р в сутки при D=50-120 мкг С/л в сутки. Как мы видим, вычисленная вероятная интенсивность поступления РО4—Р в воду в результате минерализации планктоном органического вещества достаточно хорошо соответствует измеренной с помощью изотопного метода величине его суточного потребления микро планктоном (Paf), что еще раз доказывает высокую степень сбалансированности этих процессов в экосистеме кораллового рифа.
На первом этапе изучения динамики основных биогенных элементов азота и фосфора в экосистеме рифа представлялось возможным решить эту проблему, выявив изменение содержания биогенов в воде над донными биотопами, производящими основную долю автотрофной продукции на рифе, и водной толщей, а также измерив интенсивность их потребления отдельными группами донных растений и кораллов. Такие исследования были выполнены на атолле Эниветок на стандартных трансектах, через риф-флет в зоне доминирования кораллов и в зоне доминирования макрофитов (Johannes et al., 1972; Pilson, Betzer, 1973; Webb et аl., 1975). Однако они дали пеструю и неопределенную картину изменения концентрации биогенов вдоль этих трансект. Аналогичная неопределенность направленности и абсолютных значений изменений концентрации биогенов в водах океана, проходящих над донными сообществами флета, была выявлена и другими работами (Hanson, Gundersen, 1976; Kimmerer, Walsh, 1981; Hatcher, Hatcher, 1981; Crossland, Barnes, 1983; Johannes et al., 1983). В большинстве случаев происходили отдача донными сообществами солевого и органического азота и слабое потребление минерального фосфата при отдаче его органических соединений (At kinson, 1983). В суточном балансе изменение концентрации солевого фосфата чаще всего было близким к нулю. А в таких биотопах рифа, как лагунные патч-рифы с преобладанием живых кораллов, бьта отмечена отдача солевого фосфора в суточном балансе (Andrews, Miller, 1983).
Таким образом, попытки оценить интенсивность потоков биогенов в сообществах рифа путем измерения динамики их концентрации в воде над рифом оказались в целом безуспешными (Atkinson, 1983; Hatcher, 1985), хотя и позволили сделать очень важный вывод о том, что одним из главных путей поставки азота в экосистеме рифа является процесс азотфиксации (Webb et al., 1975). Столь же неопределенные результаты дали попытки экспериментального определения динамики биогенов путем анализов изменения их концентрации в изолированных прозрачными пластиковыми или стеклянными колпаками участках дна (Henderson, 1981; Пропп, 1982, 1983; Harrison, 1983). Оказалось, что как на свету, так и в темноте концентрация солевых биогенов над дном, занятым обломочным материалом или коралловым песком, или вообще мало меняется, или чаще преобладает их слабая отдача, причем не только азота, но и фосфора.
В связи с дефицитом данных об истинной интенсивности и направленности потоков биогенов между донными биотопами рифа и водной толщей мы выполнили исследования динамики фосфата с применением изотопной метки на рифах о-ва Херон. Опыты с перифитонными обрастаниями скального флета, мертвых кораллов и крупного обломочного материала были выполнены в аквариумах. Анализы интенсивности обмена фосфора между дном и водной толщей в зоне кораллового песка производили с применением светлых и темных плексигласовых колпаков, снабженных вертунами для перемешивания в них воды после установки их на дно.
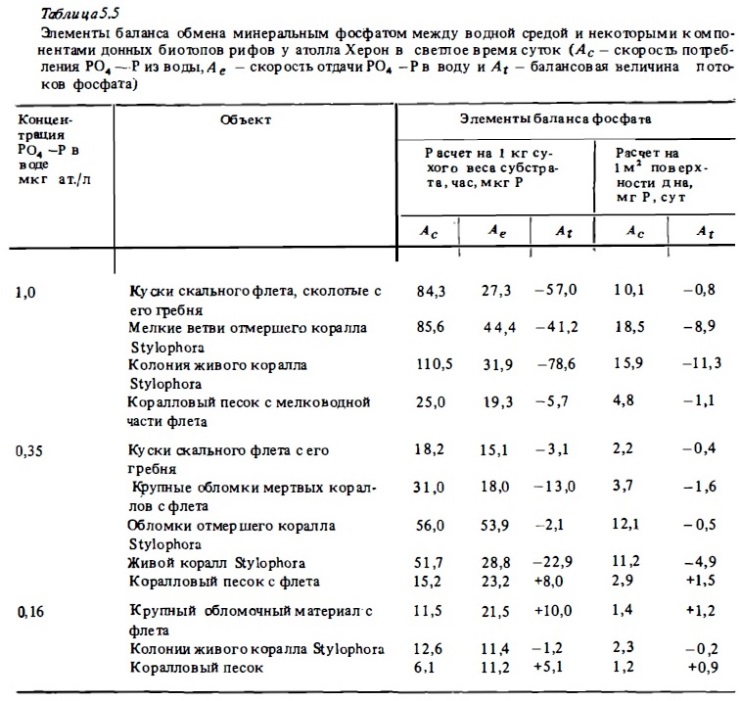
Типичные результаты определений потоков фосфата между перифитонными обрастаниями скального флета, обломочного материала и водой, выполненных в аквариумах, приводятся в табл. 5.5 и на рис. 5.4. Они показывают, что для перифитона, а также и для верхнего слоя рыхлых донных осадков рифа (кораллового песка), подобно и для самих кораллов, характерно наличие двух противоположно направленных потоков биогенов: потребления из воды и выделения. Это вполне понятно, поскольку в сообществах перифитона и в верхнем слое кораллового песка в составе слизистых структур находятся как потребляющие их автотрофы, так и выделяющие их в процессах деструкции и минерализации гетеротрофы (Сорокин, 1980). При этом, как видно из приведенных данных, общая интенсивность потоков фосфата между перифитоном и водной средой и даже соотношение интенсивностей потребления (Ас) и выделения РО4—Р весьма близки к таковым у живых кораллов (см. рис. 5.2, 5.3). В наибольшей степени показатели скорости обмена РО4—Р и его балансовых соотношений у живых кораллов . и у сообществ перифитона как расчете на сухой вес субстрата, так и в расчете на 1 м2 дна при его среднем покрытии живыми кораллами и обломочным материалом, сказываются сходными между собой (см. табл. 5.5). Существенно при этом, что интенсивность обмена (фотосинтез, дыхание) у колоний мертвых кораллов, обросших перифитоном, и у живых колоний кораллов того же вида имеет близкое значение.
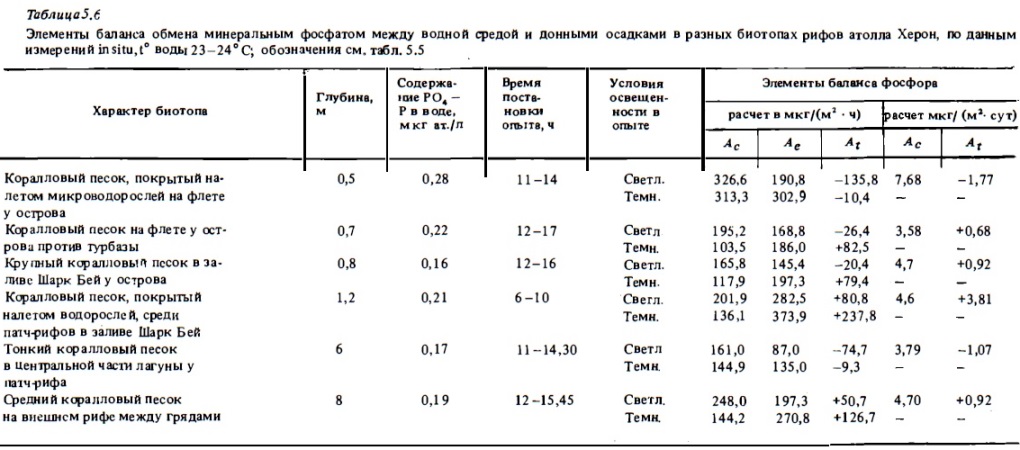
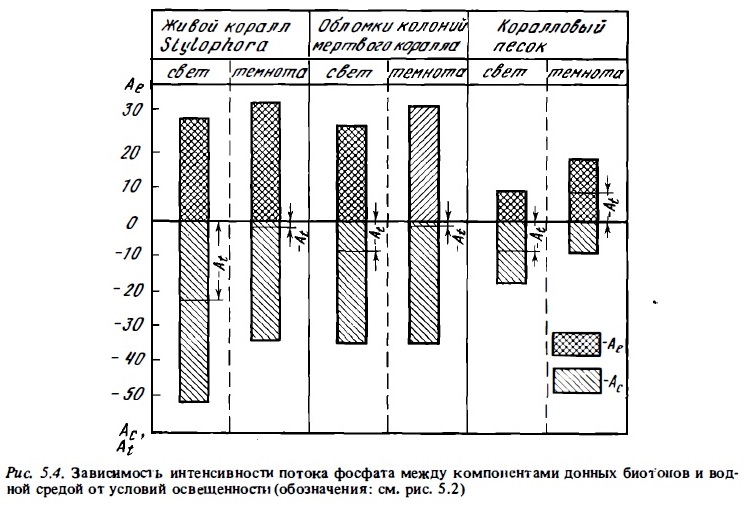
Анализ табл. 5 .5 показывает, что интенсивность потоков фосфатов между сообществом перифитона и водной средой, а также соотношением между их потреблением и отдачей в большой степени зависят от их концентрации в воде. При концентрации РО4—Р, (Ае), близкой к естественному, весьма низкому его содержанию в воде над рифами у о-ва Херон, где были выполнены нами исследования (0,1 —0,3 мкг ат/л), интенсивность потребления фосфата из воды (Ас) перифитоном скального флета и обломочного материала колеблется в пределах 10-60 мкг Р/кг сухого веса материала в час, что, как отмечалось выше, близко к таковой у живых кораллов при сопоставимой концентрации РО4—Р (см. табл. 5.3, 5.5). При таких низких концентрациях РО4—Р скорость идущего одновременно выделения перифитоном фосфатов в среду (Ае) часто была соизмеримой со скоростью его потребления (Ас). Ввиду того что она в целом в меньшей степени зависит от внешней концентрации РО4—Р, при низком содержании фосфата в воде его выделение часто превосходило его потребление. В данном случае балансовая величина At, отражающая соотношение этих процессов, имеет положительное значение, т.е. имеет место чистая отдача перифитоном фосфата (см. табл. 5.5. рис. 5.4).
При концентрациях РО4-Р 0,5-3,5 мкг ат./л интенсивность потребления РО4 -Р возрастает до 80-120 мкг Р/кг в час. При этом она значительно (в 1,5-3 раза) превосходит его выделение, которое мало повышается при росте концентрации РО4—Р. Вследствие этого имеет место чистое потребление перифитоном РО4—Р из воды. Величина At при высоких концентрациях РО4—Р в воде всегда бывает отрицательной. Эта особенность зависимости баланса фосфата и скорости его потребления из воды от концентрации на первый взгляд не может иметь существенного экологического значения, поскольку такие высокие концентрации в рифовых водах, казалось бы, вообще бывают весьма редкими. На самом же деле это, по-видимому, не так. В донных сообществах рифа обитает богатая фауна малоподвижных крупных беспозвоночных, а также рыб с территориальным поведением. Эти животные обитают постоянно в каком-либо определенном месте донного биотопа и выделяют локально значительные количества солевых биогенов, концентрация которых в зонах их выделения, часто имеющих сложный рельеф, может быть весьма значительной. Поэтому способность сообществ перифитона (а также, как мы увидим ниже, и кораллов) значительно ускорять потребление биогенов из воды при повышении их концентрации следует рассматривать как важное их свойство, которое повышает возможность использования ими биогенных ресурсов экосистемы рифа.
Рост содержания РО4—Р в воде до 2-3 мкг ат/л сопровождается почти линейным ростом скорости его потребления перифитоном (Ас) (см.рис. 5.3), а также некоторым ростом его выделения (Ае). Дальнейшее увеличение концентрации РО4—Р вызьшает перегиб кривой, которая имеет тенденцию выхода на плато при содержании РО4—Р более 9-10 мкг ат/л. Скорость выделения фосфата при этом стабилизируется, варьируя у разных типов субстратов от 30 до 70% от абсолютной скорости его потребления. Условия освещенности в целом мало влияют на общую скорость потребления РО4—Р перифитоном, а также альгофлорой и макрофлорой, населяющей коралловый песок (см. рис. 5.3, 5.4). В темноте, как и в опытах с живыми кораллами (см. рис. 5 .2), наблюдалось некоторое снижение абсолютной скорости потребления РО4—Р при увеличении скорости его выделения. Поэтому в темноте баланс сдвигался, как правило, в сторону выделения РО4—Р. Чистое его потребление (Рс) при этом снижалось, а во многих случаях в темноте величина Рс меняла знак и вместо чистого потребления наблюдалась чистая отдача РО4—Р перифитоном. Особенно часто такая ситуация наблюдалась в опытах с коралловым песком в аквариумах и in situ (табл. 5.5, 5.6).
В расчете на 1м2 интенсивность потоков фосфата (Ас и Ае) между сообществами дна рифа, занятого обломочным материалом и водной толщей, можно оценить величинами 3-10 мг Р/м2 за сутки. Аналогичные величины потоков фосфата между дюм, занятым коралловым песком, и водной толщей в соответствии с их определениями, выполненными в опытах in situ, составляют 2-7 мгР/м2 в сутки. Балансовые их величины (Лт), отражающие в чистом виде интенсивность обмена фосфатом между дном и водной толщей, оказываются в целом очень низкими, что в конечном счете и отражает полузамкнутый характер биогенов в донных биотопах рифа, о чем уже говорилось выше. В донных биотопах, занятых живыми кораллами или перифитоном, при концентрациях РО4—Р в пределах их естественного уровня (0,1 — 0,5 мкг ат/л) в среднем величинаА, близка к нулю. Это как будто означает, что биотоп практически не отдает в толщу воды солевого фосфора, несмотря на наличие интенсивных потоков РО4—Р между ним и водной толщей, которые при этом уравновешивают друг друга. Однако при оценке баланса следует учесть, что сообщества донных биотопов, занятые перифитоном и живыми кораллами, наряду с минеральным фосфатом интенсивно потребляют из водной толщи также и органические формы растворенного и взвешенного фосфора в виде растворенной органики, детрита и планктона. Эта не учтенная в описанных выше опытах приходная часть баланса фосфора и обеспечивает возможность некоторой чистой отдачи фосфата из биогенов дна в толщу воды, а также создание в донных осадках и скальных породах рифа запасов кристаллического неорганического фосфата в виде апатита и других фосфорсодержащих минералов (Entsch et аl., 1983).
В донных биотопах, занятых коралловым песком, в суточном балансе в наших опытах преобладала отдача фосфата в толщу воды в размерах 1 —3 мг/м2 в сутки при интенсивности самих его потоков между дном и водной толщей (Ас и Ае) 2-7 мг/м2 в сутки. В аналогичных опытах Харрисона (Harrison, 1983) балансовые потоки фосфата (Аг) составляли ±0,1-1,5 мг/м в сутки, меняя знак в разных участках дна лагуны. Попытка расчета соотношений между потоками фосфатов, измеренных с помощью изотопной метки, и интенсивностью метаболизма компонентов донных сообществ дает следующие результаты. При средней интенсивности дыхания и фотосинтеза в донных биотопах, занятых кораллами и обломочным материалом 2-4 г С/м2 и при величинах Ас 5-20 мг Р/м2 в сутки, отношение С:Р будет варьировать в пределах 200-400, что в 5-10 раз выше нормального их соотношения (около 40) при биосинтезе и дыхании живых организмов, не относящихся к симбионтам и не имеющих полузамкнутого обмена фосфора.
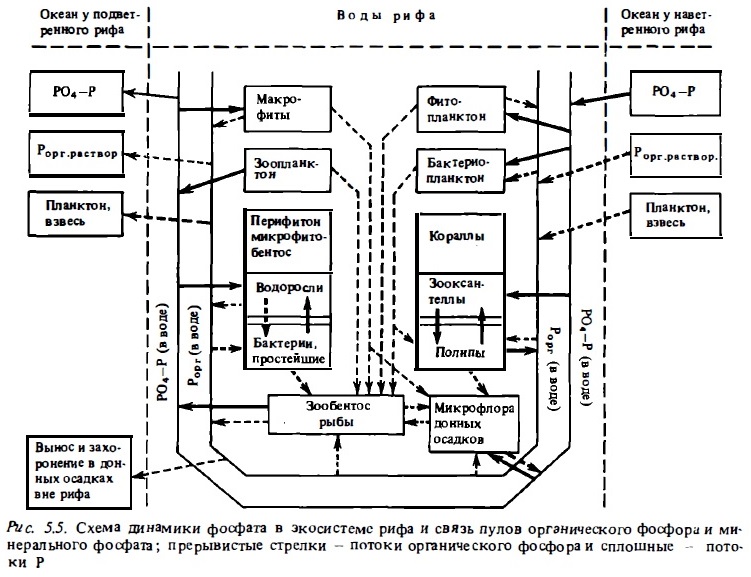
Описанная выше сбалансированность потоков биогенов между дном и водной толщей объясняет тот факт, что даже при интенсивном поступлении биогенов в экосистему рифа в условиях антропогенного загрязнения (Johannes, 1973; Kimmerer, Walsh, 1981; Crosslaid, 1983) или удобрения (Kinsey, Domm, 1974) уровень концентрации солевых биогенов в воде существенно не меняется и остается сравнительно низким, хотя при этом вследствие изменения биогенного режима происходят большие изменения в структуре его донных и планктонных сообществ и в соотношении продукционно-деструкционных процессов (Wiebe, 1985). Эти наблюдения подтверждают вывод о том, что основной мобилизуемый запас биогенов экосистемы рифа находится в составе живого вещества организмов и (в гораздо меньшем количестве) в органическом веществе детритных донных осадков (Hatcher, 1985), а также в составе запаса солевых форм и в водах, заполняющих пустоты скального тела рифа (Szamant-Froelich, 1983). Прямые измерения такого запаса не производились. Возможны, однако, ориентировочные его подсчеты, основанные на определениях общего запаса органического вещества в донных биотопах рифа (Bunt et al., 1975), который оценивается в 50-130 гС/м2, и на известных соотношениях C:N и С:Р в органическом веществе донных осадков: C:N = 12715 и С:Р = = 507100. Расчеты дают вероятные средние величины запаса биогенов в органической форме: азота 3-10 г/м2 и фосфора 0,5-2,5 г. При условии, что интенсивность потоков фосфора между водной толщей и донными биотопами составляет в среднем 3-10 мг Р (см. табл. 5.6), и азота — в среднем 40-150 мг/м2 в сутки, скорость оборота этого запаса оказывается достаточно высокой, несмотря на то что значительная часть этого запаса биогенов находится в составе биомассы крупных организмов: рыб, макрофитов, макробентоса. Среднее время оборота биогенов в донных биотопах рифа, согласно этим расчетам, должно быть близким к 2-4 мес. Результаты исследований динамики фосфата обобщены на схеме, рис.5.5.