
Первая серьезная попытка создания математической модели на основе трофодинамических отношений компонентов рифа была сделана большим коллективом ученых (более 40 человек) в рамках проекта CITRE. Группа поставила перед собой задачу выделить основные функциональные компоненты экосистемы, объединить их в блоки-сообщества, составляющие трофические уровни (Dahl et al., 1974). Уже на стадии этого выделения оказалось 104 компонента, между которыми могло существовать более 10000 отдельных взаимодействий. Более того, их объединяли более 2000 реальных трофических и прочих взаимосвязей. Вполне очевидно, что такая модель из-за трудностей на практике реализовала не была.
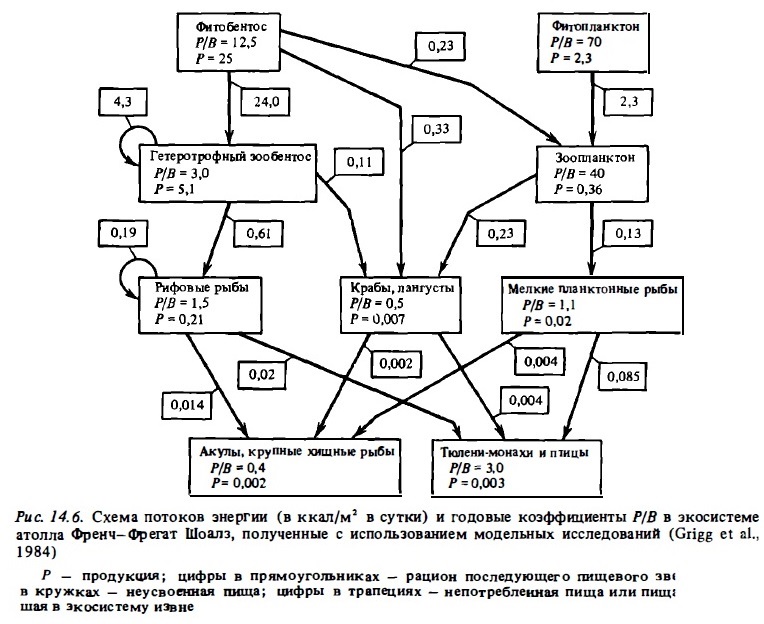
В 1980-1982 гг. была сделана попытка создать математическую модель экосистемы рифа в рамках проекта ЕСОРАТН Гавайского университета. Модель разрабатывалась для конкретного рифа одного из гавайских атоллов — Френч Фрегат (Polovina, 1984; Atkinson, Grigg, 1984; Grigg et al., 1984). Это большой серповидный риф с мелководной лагуной. Он занимает площадь около 700 км2. За основу для построения модели была принята упрощенная схема трофической сети рифа. Схема содержала органическое число компонентов - всего 13 вместо 104, хотя в нее были включены некоторые малозначимые компоненты, например черепахи. Экосистема этого атолла рассматривалась в изоляции от экосистемы океана, вернее, учитывалась только отдача вещества рифом и не учитывалось поступление планктона и взвеси из вод океана. Кораллы были объединены с донными водорослями. Из пищевой сети и трофических связей были полностью исключены детрит и РОВ. В единые компоненты были объединены зоопланктон хищный и мирный, а также зообентос и рифовые рыбы. Не был учтен демерсальный характер зоопланктона рифа и его использование зообентосом и донными рыбами. Весь полевой материал, собранный на атолле Френч Фрегат и составивший фундамент модели, состоял из результатов выборочных анализов состава пищи крупных хищных рыб (акул, каранксов), а также из приблизительных оценок запасов (биомассы) хищников конечного звена: рыб, тюленей, птиц. Остальные сведения, необходимые для построения модели, и расчетные коэффициенты бьши взяты из литературы и относились преимущественно к сообществам и организмам умеренных вод.
Главным итогам расчетов с использованием модели была величина энергетического "входа" — первичная продукция. Она оказалась равной 4,3·106 кг/км2 сырой биомассы в год или около 1,2 г С/м2 в сутки. По ней и производилась верификация модели путем измерения первичной продукции донных сообществ в отдельных зонах атолла Френч Фрегат в разные сезоны года (Grigg et аl., 1984). Они дали близкую к указанной выше модельной величине годовую продукцию 6,1·106 кг/км2 в год сырой биомассы. Главным результатом модельных расчетов явилась схема энергетики экосистемы атолла Френч Фрегат, в которой рационы и продукция была выражены в килограммах сырой биомассы на квадратный метр в год. Чтобы сделать ее более сопоставимой с приводимыми нами схемами, мы изменили в ней величины продукции в ккал/м2 в сутки (рис. 14.6).
Таким образом, упрощенная, правда, содержащая благодаря этому ряд неверных предпосылок модель ЕСОРАТН была успешно верифицирована. Авторы рекомендуют использовать эту модель для разработки стратегии управления ресурсами рифа. Успех этой модели объясняется подбором подходящих коэффициентов К1, Р/В и смертности (выедания). Например, заложив в модель заведомо завышенный коэффициент выедания (0,85), авторы с помощью модели приходят к выводу о том, что именно выедание (а, скажем, не биогены) контролирует продукцию экосистемы. Правда, вряд ли было обоснованным полагать, что в сообществах. рифа с его обилием убежищ выедание может быть выше, чем в пелагиали, где его принимают близким к 0,6-0,7. Однако, взяв для своей модели величину выедания 0,5, авторы получили абсурдно высокую величину первичной продукции — более 200·106 кг/км2 в год, что в 30 раз выше той, которая была получена ими при полевых измерениях. Эти свойства модели вполне объяснимы заниженными коэффициентами К1 (в среднем 0,17) и отсутствием звена бактерий. Отсутствует в модели и детритная пшцевая цель, столь важная в сообществах рифа (Peres, Picard, 1969; Gerber, Marshal, 1974). В реальной открытой системе рифа нет внутренней сбалансированиости, принятой в рассматриваемой модели. Биофильтр экосистемы рифа потребляет значительную долю энергии из проходящих над ним вод океана. Итак, упрощение экосистемы рифа, к которому прибегли авторы модели ЕСОРАТН, сделав реальным ее создание, лишило ее адекватности, и, следовательно, снизило ее научную ценность. Хотя всем понятно, что всякая модель является всего лишь гипотезой, она не должна игнорировать уже известные факты.
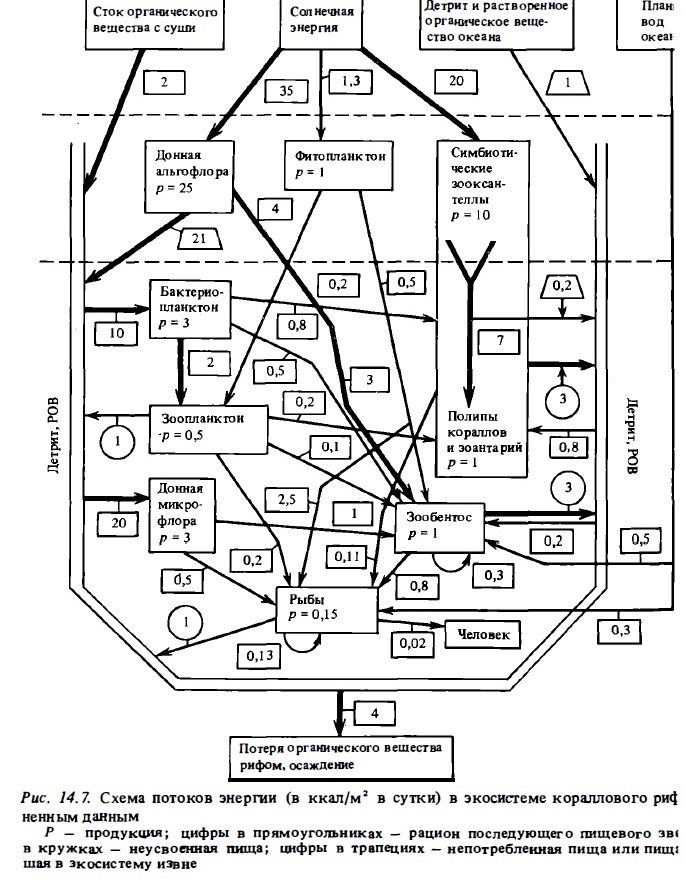
Энергетический вход в экосистему рифа формируется прежде всего за счет фотосинтеза донных растений: макрофитов, кораллин, микрофитобентоса, морских трав, а также зооксантелл симбионтов: кораллов, фораминифер, тридакн. Дополнительными источниками внешней энергии являются фотосинтез фитопланктона, а также планктон, взвесь и РОВ вод океана, проходящих над рифом. Первые два компонента поставляются в экосистему рифа путем их потребления организмами биофильтра рифа рыбами-планктофагами, а последний (РОВ) преимущественно за счет его потребления перифитонной микрофлорой. На барьерных и окаймляющих рифах существенным источником внешней органики служит ее приток с суши и из прибрежных мангров. Это органическое вещество включается в пищевую цепь через детритный пул при участии бактерий. Значительная часть продукции фитобентоса не выедается и также включается в пищевую цепь через детритный пул. Поток энергии от донной альгофлоры через детритный пул к фауне является главным путем использования продукции автотрофов в гетерогенных процессах, протекающих в экосистеме рифа. Второй по величине поток энергии идет от зооксантелл к тканям животных-хозяев. Этот поток идет преимущественно внутри их тела. Однако последние экскретируют до 30% этой транслоцированной продукции в виде слизи и РОВ, которые пополняют детритный пул и через него включаются в бактериальное звено. Бактерийное население использует для своего метаболизма около 70% всего потока энергии в экосистеме рифа (вспомним, что в модельной схеме ЕСОРАТН, рис. 14.6, этот компонент экосистемы вообще отсутствует). Важным путем использования донной растительности рифа является также и прямое выедание ее рыбами, ежами и гастроподами.