
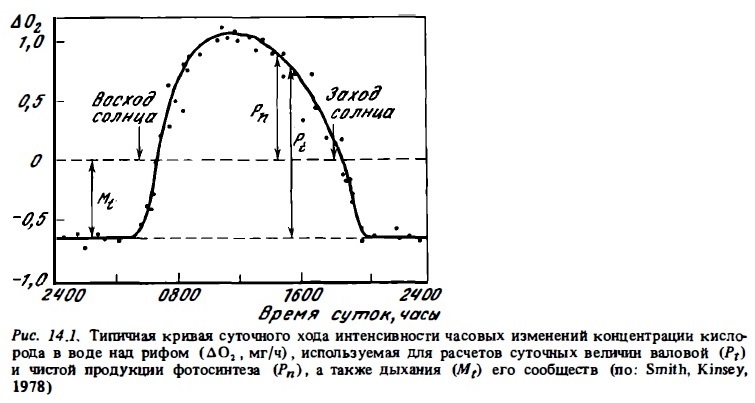
В биогеохимическом метаболизме рифа доминируют процессы фотосинтеза, дыхания и кальцификации. Их интенсивность выражают обычно в граммах углерода на квадратный метр поверхности дна. В рыхлых донных осадках лагуны, каналов, подножия рифа существенное значение имеют также и анаэробные процессы, среди которых преобладают сульфатредукция и денитрификация. Уже первые исследования биогеохимического метаболизма на тихоокеанских атодлах показали, что коралловые рифы — это автотрофные по балансу экосистемы, которые функционируют при высоком уровне продуктивности и при высоких скоростях обмена в обедненной солевыми биогенами среде (Odum, Odum, 1955). Однако на рифах, расположенных вблизи больших островов и получающих значительный приток органического вещества с суши, в балансе они часто бывают гетеротрофными (Gordon, Kelley, 1962; Coreau et аl., 1971; Kinsey, 1985). Сама сбалансированность автотрофных и гетеротрофных. процессов в экосистеме рифа на биогеохимическом уровне выражается в соизмеримости валовой продукции органического вещества всеми растениями и симбиотическими водорослями (Pt) и гетеротрофной его деструкции в процессах дыхания всех гетеротрофой рифа (Mt), где Рt:Мt~1 (Kinsey, 1983, 1983а) ). В отдельных биотопах, в зонах или фациях рифа это соотношение может быть больше или меньше единицы (Сорокин, 1980а; Жариков, 1987), но в целом в пределах экосистемы рифа как единого целого оно обычно близко к единице или (в условиях значительного терригенного стока) несколько меньше. Сбалансированность автотрофных и гетеротрофных процессов на рифе указывает на то, что чистая продукция сообщества рифа (Рn) очень мала, поскольку она равна разности Pt—Mt. Тем не менее ее определение имеет существенное значение для оценки возможных масштабов в эксплуатации ресурсов рифа. Действительно, изъятие вторичной продукции без разрушения системы возможно лишь за счет ресурсов чистой продукции.
Определение размеров чистой продукции осложняется тем, что при рассмотрении энергетического баланса экосистемы рифа обычно учитывают как приходную часть только первичную продукцию донных биотопов рифа, игнорируя первичную продукцию фитопланктона и другие вероятные ее источники, роль которых на многих рифах может быть соизмеримой со значением фотосинтеза донных автотрофов. В их числе поступление органического вещества с терригенным стоком, нагоны синезеленых водорослей, постоянное поступление в зону рифа с проходящими над ним водами океана зоопланктона, выедаемого рифовьыми рыбами и кораллами, поступление с водами океана растворенного органического вещества и детрита, которые используются в качестве пищи донными фильтраторами рифа и включаются в пищевую сеть. Что касается расходной части баланса, то наряду с тратами на обмен сообщества рифа теряют значительное количество энергии в виде органического вещества и планктона, которые выносятся с рифа в океан с уходящими с него водами. Эти источники должны, безусловно, учитываться при анализе энергетического баланса экосистемы рифа. Как отмечалось выше, в пределах отдельных биотопов рифа (лагуна, флет, внешний склон) отношение Pt/Mt может сильно варьировать. В одних биотопах рифа (флет, внешний склон) обычно преобладают автотрофные процессы фотосинтеза, в других, таких, как лагуна, зона рва ("moat") у внешнего края рифа, преобладают процессы гетеротрофной деструкции, питаемые за счет поступления в эти биотопы органического вещества из зон, где преобладают автотрофные процессы. Вполне очевидно также, что преобладание гетеротрофных деструкционных процессов над автотрофными неизбежно в зонах рифа, куда в большом количестве поступает органическое вещество с терригенным стоком.
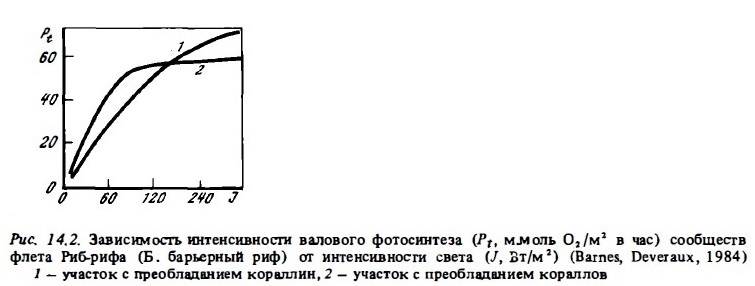
Сам уровень автотрофной продукции большей части донных биотопов коралловых рифов, да и экосистем коралловых рифов в целом, оцениваемый по величинам их суточного валового фотосинтеза, оказался близок к рекордному его уровню (8-15 г С/м2 в сутки), который характерен лишь дня экосистем тропических лесов, зарослей макрофитов на шельфе умеренных морей (келп) или зон интенсивного апвеллинга в океане. Столь высокие величины первичной продукции возможны лишь при многоярусном ми объемном (ветвистом), но не однослойном распределении фотосинтезирующих элементов (листьев, клеток фитоцианктона ми зооксантелл) в пределах освещенной зоны (Odum et аl., 1958; Dabl, 1976; Jokiel, Morrissey, 1986). Основные фотосинтезирующие агенты рифа — кораллы, макрофиты и морские травы создают ветвистую многоярусную структуру, благодаря которой суммарная фотосинтезирующая поверхность биотопов рифа в несколько раз (от 5 до 25 раз) превосходит проективную площадь дна (табл. 14.1). Такое возрастание фотосинтезирующей поверхности в сочетании со способностью фотосинтетического аппарата кораллов и макрофитов, обитающих в условиях недостатка света к световой адаптации, обеспечивает высокий квантовый выход фотосинтезирующих донных сообществ и в конечном итоге высокую первичную продукцию (Beyers, 1966). Существенно, что порог светового насыщения в сообществах флета, как правило, бывает ниже максимальной интенсивности солнечной радиации в середине дня в 1,52 раза и составляет от 100 до 200 Вт/м2, несмотря на частичную затененность поверхности фотосинтезирующих организмов (рис. 14.2). Это позволяет им функционировать с высокой эффективностью в течение большей части светового дня. Как видно на рис. 14.1, в период с 8 ч утра до 16 ч фотосинтез превышает в них дыхание.
Формирование представлений о биогеохиминеском метаболизме рифа и его механизмах идет как путем его изучения в целостным сообществах, так и путем накопления и обобщения данным о функциональной активности отдельным ее компонентов. При этом большинство исследователей в числе компонентов рифа не учитывают планктон как таковой, считая, что его вклад в общий метаболизм рифа очень мал. Надо определения показали некорректность такого подхода. Вклад планктонного сообщества рифа в его суммарный метаболизм и особенно в процессы гетеротрофной деструкции (дыхания) весьма существен и составляет 10-30% от его суммарной величины. В числе основным компонентов донным биотопов рифа, создающих автотрофную продукцию; зооксантеллы симбионтным животным (кораллов, эоантарий, фораминифер), донные водоросли (талломные макрофиты, нитчатки, микроводоросли перифитона и песка, кораллины, эндолитические синезеленые водоросли, а также морские травы) (Lewis, 1977).
Вопросы методологии изучения биогеохимического метаболизма в экосистеме рифа и основные результаты исследований по данной проблеме обсуждены и обобщены в ряде обзоров (Lewis, 1977, 1981; Sournia, 1977; Kinsey, 1983, 1983а, 1985).
