Для решения вопроса об энергетическом балансе у кораллов были применены разные подхода. Мы попытались решить его, определив долю разных источников гетеротрофного питания в суточном балансе энергии, приняв, что траты на обмен (Mt) составляют 60% от общих энергетических потребностей. Схема расчетов доли гетеротрофного питания в общем балансе энергии коралла дана выше. В соответствии с расчетами, основанными на результатах экспериментальный определений интенсивности гетеротрофного питания кораллов в конкретных условиях освещенной зоны рифа у о-ва Херон, сообщество герматипных кораллов формирует свой энергетический баланс следующим образом: 70% энергии дает фотосинтез зооксантелл, 20% — хищное питание и 20% — питание бактериопланктоном и РОВ. Согласно аналогичным расчетам Люиса (Lewis, 1981), зооксантеллы обеспечивают в среднем 65% общих энергетических потребностей герматипных кораллов.
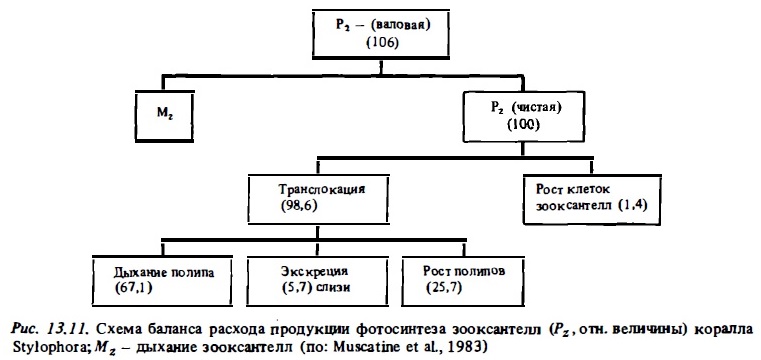
Другой подход к решению этого вопроса состоит в определении величин чистой продукции зооксантелл (Рnz), процента транслокации продукции зооксантелл в клетки полипа-хозяина (T), дыхание самого полипа за вычетом дыхания зооксантелл (Ма). Тогда доля автотрофного питания полипа в компенсации своих трат на обмен (CZAR) будет выглядеть следующим образом: CZAR = Pnz·Т/Ма% (Muscatine et al., 1984, 1985).
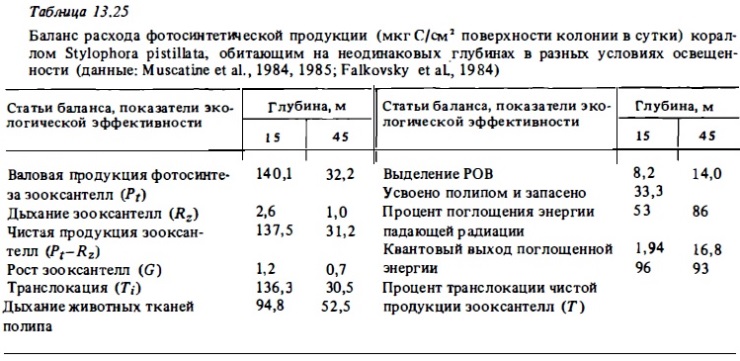
Абсолютная величина транслокации (Ti) для этих расчетов была получена как разность между чистой продукцией зооксантелл и их тратами на их собственный рост (G). Тогда Т = (Pnz-G)·100%/Pnz. Удельная скорость роста зооксантелл определялась путем измерения времени их генерации по частоте встречаемости во взвеси свежевыделенных зооксантелл и клеток, ядро которых находилось в стадии метоза (Wilkerson et аl., 1983). В результате упомянутыми авторами был вычислен баланс расхода энергии фотосинтетической продукции колониями коралла (Stylophora pistillata, обитавшими на глубинах 15 и 45 м при разных условиях освещенности (табл. 13.25, рис. 13.11). Как видно из этой таблицы, колонии кореша Stylophora в освещенной зоне способны полностью компенсировать все свои энергетические траты только за счет фотосинтеза зооксантелл, тогда как колонии, живущие при недостатке света, могут компенсировать за счет фотосинтеза менее половины своих энергетических потребностей, хотя отношение Р:М у них в дневное время выше 1. Эти колонии получают недостающую энергию за счет гетеротрофного питания. Величина транслокации продукции фотосинтеза зооксантемами, согласно балансовым расчетам, должна составлять у этого коралла около 98%.
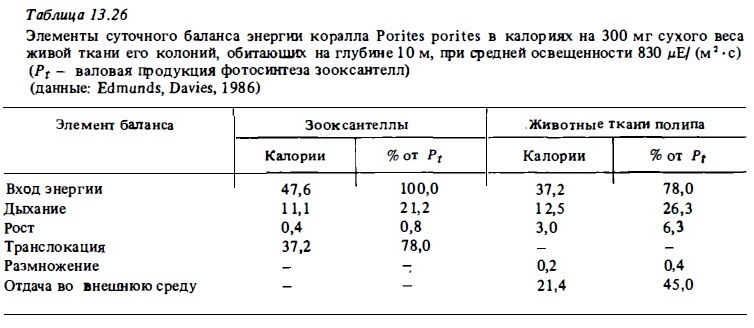
Аналогичные определения баланса энергии были выполнены для колоний кораллов Pocillopora eydouxi (Dawis, 1984) и Porites porites (Edmund, Davis, 1986). Однако в этих случаях авторы определяли дыхание самих зооксантелл во взвеси in vitro и получили их величины от 9 до 21% от валовой продукции фотосинтеза вместо 2-3%, полученных расчетных методом. Соответственно последние авторы определили для перечисленных выше кораллов меньшие величины транслокации: 90% для колоний коралла Pocillopora и 78% для коралла Porites (табл. 13.26). Сами полипы расходуют на дыхание около трети транслоцированной продукции фотосинтеза. Остальную энергию они тратят на выделение слизи и создание запасных веществ.
Описанные попытки анализа баланса энергии кораллов, безусловно, являются первым приближением к решению проблемы и не лишены серьезных недостатков. В частности, в балансе коралла Stylophora, представленном в табл. 13.25, явно занижены расчетные траты зооксантелл на обмен: у свободно живущих водорослей они близки к 15-20% от продукции фотосинтеза. Явно занижена и измеренная в кратковременных опытах: с помощью С1 4 отдача кораллами органического вещества. В этом случае отдача его происходит в основном за счет немеченого вещества, которое с помощью С1 4 не учитывается. По нашим прямым измерениям эта отдача близка к 20% от валового фотосинтеза коралла в освещенном биотопе рифа, а не 5%, как это следует из табл. 13.25. Поэтому и процент транслокации (95%) несомненно этими авторами завышен. Его реальную величину можно оценить в 70-80% от Pt. В соответствии с измерениями Титлянова и др. (1988в) процент транслокации (точнее: CZAR-процент транслоцированной продукции в дыхании животной ткани полипа) близок к 70-75%. С этой точки зрения более реален баланс энергии коралла Porites porites, представленный в табл. 13.26.
Удельная скорость роста зооксантелл кораллов (суточный Р/В-коэффициент) оценивается величинами 0,009-0,013 за сутки (время одной генерации 40-70 дней (Wilkerson et аl., 1983; Muscatine et аl., 1984, 1985). Те же авторы оценивают удельную скорость роста полипа коралла Stylophora по данным прямых измерений близкой к 0,002-0,004 в сутки или 0,7-1,5 генераций в год. Близкие величины удельной скорости роста живых тканей полипов (около 0,003 в сутки) были получены для коралла Porites (Edmunds, Davies, 1986). При такой же величине время удвоения массы живой ткани полипов будет близким к 9 мес. Согласно же Вилкерсону и др. (Wilkerson et аl., 1983), удельная скорость роста зооксантелл у коралла Montastrea значительна (0,01-0,4 в сутки), что намного выше скорости роста тканей самого полипа. Тем не менее полшы могут регулировать численность зооксантелл в тканях путем частичного переваривания их избытка (Reimer, 1971; Fitt, Trench, 1980; Colley, Trench, 1985). Мы выполнили расчеты удельного прироста (G) биомассы полипов массовых видов кораллов, заселяющих верхние освещенные горизонты рифов, пользуясь физиологическим методом расчета трат энергии на рост:
G = МtК₂/1-К₂,
где К₂ - коэффициент использования усвоенной пищи на прирост и Mt — траты на дыхание.
Для разных видов кораллов величина суточных коэффициентов Р/В (в данном случае - G/B) варьировала в пределах 0,006-0,02 (Сорокин, 1984в). Согласно сходным расчетам Люиса (Lewis, 1981), в среднем для сообщества герматипных кораллов величина суточного коэффициента Р/В близка к 0,005 (годовой коэффициент Р/В — 1,8-2,0). Несомненно, что такие относительно небольшие траты энергии на рост полипов с избытком обеспечиваются энергией за счет транслокации и гетеротрофного питания. Поэтому скорость роста кораллов лимитируется не ею, а интенсивностью кальцификации и ограниченной скоростью роста известкового скелета, а также (в меньшей степени) недостатком связанного азота, поскольку в транслоцируемых зооксантеллами продуктах фотосинтеза преобладают безазотистые соединения (отношение C/N в них ~ 30, Muscatine et аl., 1984).