Зооксантеллы кораллов, которые обитают в верхних освещенных зонах рифов, адаптированы к высокому уровню освещенности. Они функционируют при слабом свете намного хуже, чем зооксантеллы кораллов того же вида, обитающих в затененных или глубинных биотопах с низкой освещенностью (Dustan, 1982). Механизм адаптации самих зооксантелл к пониженной освещенности состоит главным образом в увеличении размеров фотосинтетических единиц (PSU). Фотосинтетические единицы представляют собой белково-пигментные комплексы, которые усваивают световую энергию и передают ее реакционным центрам фотосистем 1 и 11. Средний размер (PSU) в зооксантеллах кораллов, адаптированных к слабому свету, почти в 1,5 раза выше, чем у кораллов из верхних освещенных биотопов (Leletkin, Zvalinski, 1981).
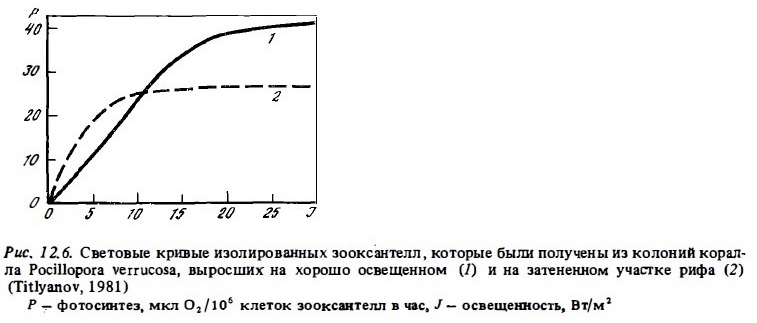
Световые кривые изолированных зооксантелл заметно отличаются от световых кривых целых колоний кораллов (Звалинский, 1978; Zvalinski et al., 1980). Главное их различие состоит в том, что они выходят на плато при освещенности 20-25 Вт/м2 (0,07-0,15 Е/м2 в час). Это в 3-5 раз ниже порога светового насыщения в опытах с целыми колониями кораллов (рис. 12.6). Другое их отличие состоит в том, что при высокой интенсивности света изолированные зооксантеллы проявляют световое угнетение. Судя по характеру их световых кривых, они имеют более низкий порог светового насыщения даже при сравнении их со свободно живущим фитоднанктоном (Scott, Jitts, 1977). Зооксантеллы, выделенные из кораллов, обитающих в затененных зонах рифа, проявляют повышенную чувствительность к слабому свету. Световые кривые зооксантелл таких адаптированных кораллов выходят на плато при освещенности всего 10-15 Вт/м2 (см. рис. 12.6).
Интенсивность фотосинтеза зооксантелл в среднем близка к таковой фитопланктона. Так, часовое ассимиляционное число у них варьирует в пределах 1-5 мг С/мг хлорофилла "а" (Wethey, Porter, 1976; Falkovsky, Duvinsky, 1981). У фитопланткона оно варьирует от 1 до 10 (Scott, Jitts, 1977).
Абсолютные величины интенсивности фотосинтеза зооксантелл варьируют в широких пределах. Они могут варьировать, по данным разных авторов, от 6 до 10 мкг О2/106 клеток в час (Zvalinski et al., 1980; Muscatine, 1980; Титлянов и др., 1988). Такой большой диапазон этих величин связан в значительной степени с различиями в методике измерений. Наиболее реальны величины 10-20 мкг О2/106 клеток В час. Дькание самих зооксантелл составляет, по разным данным, от 1% (Muscatrne et al., 1984) до 20% (Zvalinski et al., 1980; Davies, 1984) от фотосинтеза. При этом низкие величины днгхания зооксантелл были получены расчетным методом исходя из соотношения биомасс (по белку) зооксантелл и всего полипа (Falkovsky et al., 1984), а более высокие — путем прямых измерений днгхания изолированных зооксантелл. В последнем случае они выражались величинами 3-5 мкг О2/106 клеток в час (Edmunds, Davies, 1986). В соответствии с результатами измерений дыхания изолированных зооксантелл кораллов полагают, что отношение валового фотосинтеза зооксантелл к их дыханию (Рt/Мz) составляет 5-11.
Суточные величины коэффициентов Р/В у зооксантелл, как правило, выше, чем у свободного живущего фитопланктона. Они составляют у них в среднем 1,5-5 за сутки (Сорокин 19846; Титлянов и др., 1988). При этом удельная скорость роста самих зооксантелл очень мала и оценивается величинами 0,003-0,01 в сутки (Muscatine, 1983; Wilkerson et al., 1983; Edmunds, Davies, 1986). Траты на собственные нужды крайне низкие — не более 10-15%. Основная часть продуцируемого ими органического вещества передается полипу-хозяину.
Что касается биогенного питания зооксантелл, то они могут использовать не только биогены, которые образуются в клетках полипа в результате минерализации (Yonge, Nicholls, 1931; Muscatine et al., 1984), но и солевые биогены, потребляемые герматипными кораллами из внешней среды (Kawaguti, 1953; Muscatine, Porter, 1977; Webb, Wiebe, 1978). Изолированные зооксантеллы в опытных условиях интенсивно потребляют солевые биогены (D’Elia et al., 1983). При этом 70-90% продуцируемых зооксантеллами органических соединений азота и фосфора транслоцируется в клетки полипа-хозяина (Muscatine et al., 1984). Именно таким путем обеспечивается синтез большей части аминокислот, необходимых полипам для их роста (Burris, 1983; Wafar et al., 1985).
Асиммиляция СО2 при фотосинтезе зооксантелл кораллов преимущественно идет через цикл "С3" с участием рибулезодифосфат карбоксилазы (Benson et al., 1978). Не исключено частичное включение также и цикла "С4" с участием фосфоэнолцируват-карбоксилазы (Trench, 1979; Trench, Fisher, 1983). Предполагается также, что зооксантеллы, находясь в клетках хозяина, имеют высокий уровень фотодахания, что может существенно увеличивать их чистую продукцию (Muscatine, 1980). Спектр первичных продуктов фотосинтеза зооксантелл, определенный с помощью С1 4, достаточно широк. Он включает углеводы, фосфоглицериновую кислоту, ряд простых аминокислот (серин, аланин, глицин, глютаминовую кислоту), а также разнообразные линиды (Von Holt, Von Holt, 1968; Trench, 1971, 1974; Schmitz, Kremer, 1977; Patton et al., 1977). Основная часть углеводов, продуцируемых зооксантеллами, представлена глюкозой и глицерином (Muscatine, Cernichari, 1969; Trench, 1974), иногда маннозой (Schmitz, Kremer, 1977). Согласно последним авторам, до 50% ассимилятов зооксантелл быстро превращаются в спирторастворимую липидную фракцию.