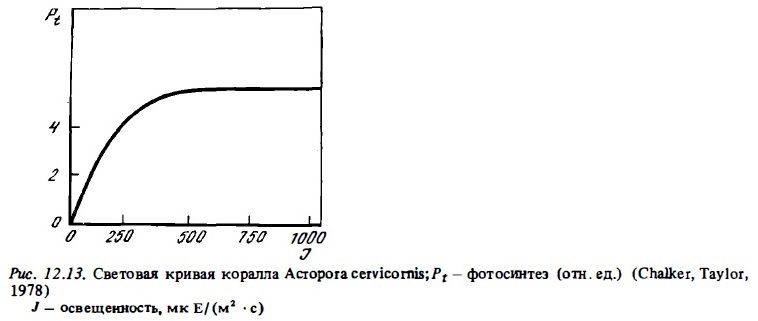
Таким образом, по-мере снижения освещенности в месте обитания кораллов увеличиваются потенциальные возможности их фотосинтетического аппарата, что проявляется в повышении уровня валового и чистого фотосинтеза в расчете на единицу площади поверхности колонии, измеряемого при постоянной умеренной освещенности (Zvalinsky et аl., 1980; Titlyanov, 1981; Falkovsky, Dubinsky, 1981). Рост потенциального фотосинтеза у адаптированных колоний непосредственно не связан с увеличением концентрации фотосинтетических единиц в тилакоидах хлоропластов их зооксантелл, как у некоторых водорослей (Лелеткин и др., 1980), а обеспечивается главным образом ростом численности зооксантелл и увеличением размера их фотосинтетических единиц.
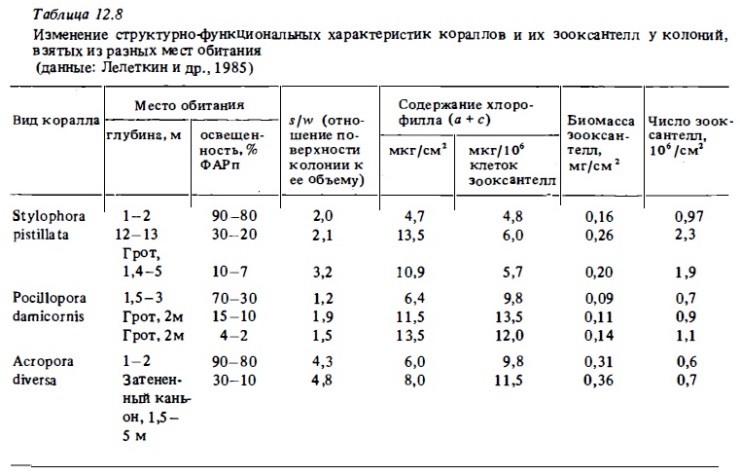
Как уже отмечалось выше, герматипные кораллы населяют очень широкий диапазон глубин (0-145 м) с диапазоном освещенности около 4 порядков (от 300 до 0,1 Вт/м2) и с очень широким диапазоном спектрального состава: от дневного и поверхности до сине-зеленого на большой глубине (Schuhmacher, Fricke, 1984). Исследования показали наличие у кораллов системы адаптации к изменениям освещенности и спектрального состава света. Среди них — изменение числа полипов и биомассы зооксантелл, а также рост содержания хлорофилла в клетках (Titlyanov et al., 1980; Dustan, 1982; МсCloskey, Muscatine, 1984; Dubinsky et al., 1984; Falkovsky et al., 1984; Porter еt al., 1984), снижение интенсивности дыхания (Davies, 1977), изменение морфологии колоний (Титлянов, 1987) и появление специальных органов и структур, повышающих возможность использования колониями слабого света (Vereshi, Fricke, 1986). Нижний предел освещенности, при котором могут обитать герматипные склерактинии, варьирует для разных видов кораллов в зваисимости от их способности к световой адаптации от 10% ФАРп (15-25 Вт/м2 у акропор) до 3-1% ФАРп у поциллопор, стилофор и турбинарий (Falkovsky-Dubinsky, 1981; Falkovsky et аl., 1984; Лелеткин и др., 1985; Титлянов и др., 1985). При этом у таких кораллов с выраженной способностью к световой адаптации даже при сильном затенении (7-2% ФАРп, или 15-5 Вт/м2) валовая продукция снижается всего на 40-50%, тогда как у макрофитов и фитонианктона при таком затенении дна снижается в 5-10 раз. Отношение валового фотосинтеза к дыханию в суточном цикле (Pt : Mt) остается выше 1 до уровня освещенности 2010% ФАРп. Валовой фотосинтез таких кораллов при освещенности 10% ФАРп всего в 1,1-1,4 ниже такового в условиях оптимального светового потока (Taylor, 1973; Jaubert, 1977). При этом кораллы, которые обитают в затененных участках мелководий (в нишах, в гротах), способны выживать при освещенности в 2-3 раза меньшей (1,5-3% ФАРп), чем кораллы тех же видов, которые заселяют открытые участки глубинных зон рифа, где недостаток света вызван его поглощением в толще вода. На глубинных участках внешнего склона рифа кораллы Pocillopora damicornis и Р. verrucosa выживают только при освещенности 7-10% ФАРп (Титлянов и др., 1985б). Выживание кораллов при меньшей освещенности в первом случае связано, несомненно, с попаданием в ниши и гроты дополнительных порций света отраженного от солнечных бликов.
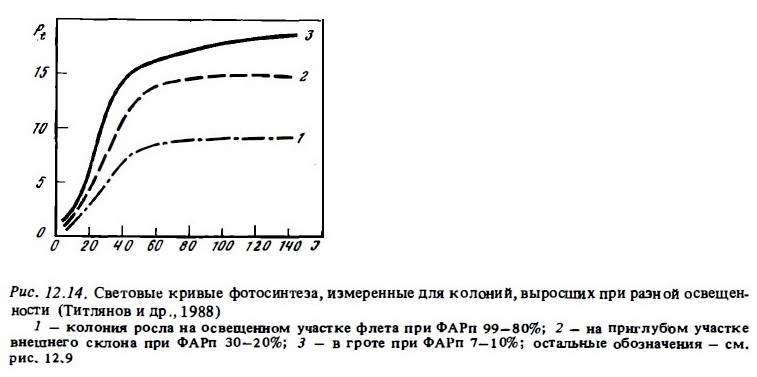
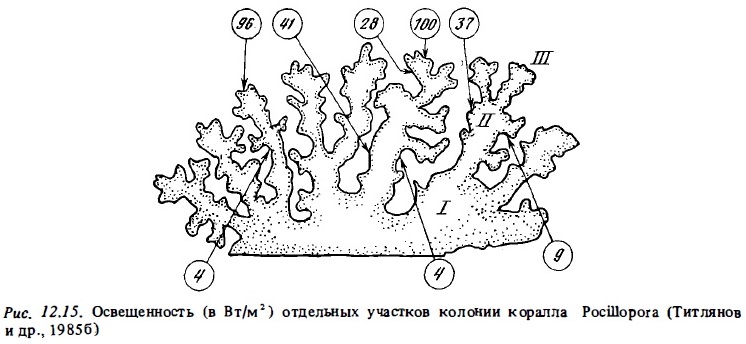
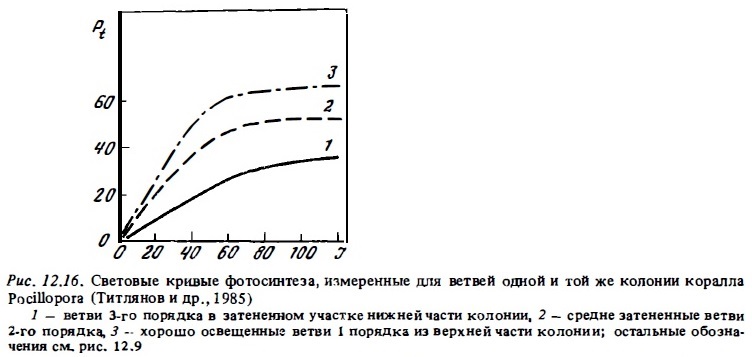
Существенно также, что свойствами адаптированности к пониженной освещенности, как оказалось, обладают затененные участки колоний ветвистых кораллов, обитающих в освещенной зоне рифа (Титлянов и др., 1985б). Измеренная с помощью специального микрозонда освещенность разных участков ветвей 1, 2 и 3-го порядков у колоний коралла Pocillopora, растущих на освещенном мелководье, варьировала в пределах 100-50% ФАРп на верхних поверхностях ветвей 3-го порядка до 9-4% ФАРп на нижних участках ветвей 1-го порядка (рис. 12.15). Ветви первого порядка, которые находятся в значительном затенении даже при наружном освещении 150-200 Вт/м2, проявляют в опытах все признаки световой адаптации, свойственные колониям, постоянно обитающим в условиях затенения. Их световые кривые имеют большой угол наклона. Они характеризуются вдвое большим потенциальным фотосинтезом при насыщающей освещенности (рис. 12.16) и почти вдвое меньшей интенсивностью дыхания (Титлянов и др., 19856). Вследствие такой световой адаптации полипов затененных участков колоний коралла Pocillopora, обитающих на освещенном мелководье, интенсивность их фотосинтеза, измеренная с помощью С1 4, была в среднем всего в 1,2 раза ниже, чем у полипов с хорошо освещенных верхних ветвей 3-го порядка. У кораллов, которые обитают на слабо освещенных участках, эта разница составляет около 3 раз. В то же время благодаря адаптивным изменениям формы колоний по мере убывания освещенности в местах обитания происходит снижение степени их самозатенения. В результате этого при одинаковой освещенности самозатенение выше у колоний из освещенных биотопов рифа (Титлянов, 1987).
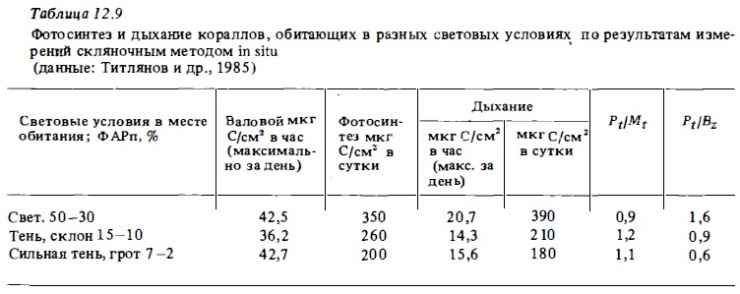
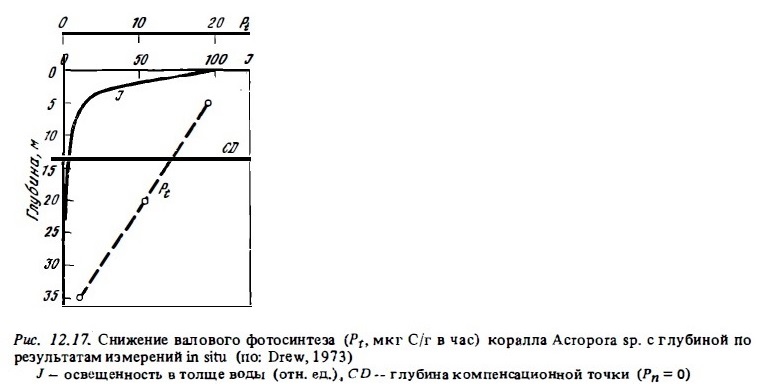
В целом наибольшая валовая продукция фотосинтеза in situ в расчете на площадь поверхности колоний или на их вес отмечается у кораллов, обитающих при умеренной освещенности 20-40% ФАРп. Кривая изменения интенсивности фотосинтеза коралла с глубиной, по данном измерений in situ, представлена на рис. 12.17. Как видно из графика, отношение Pt : Mt у подопытного коралла Acropora sp. было выше 1 до глубины 14 м, на которой находилась компенсационная точка, где валовой фотосинтез был равен дыханию. Освещенность же на этой глубине составляла около 1-2% ФАРп. С применением С1 4 было показано, что у колоний этого коралла, живущих в глубинной зоне внешнего склона рифа при малой интенсивности света, фотосинтез обнаруживается до глубины 35 м, где освещенность составляет доли процента от наружной (Drew, 1973). В прозрачных же водах Красного моря компенсационная точка фотосинтеза кораллов находилась на глубинах 80-100 м (Fricke, Schuhmacher, 1983).
Как мы видели выше, многие видах кораллов способны заселять биотопы рифа в широком диапазоне освещенности при различии ее крайних значений на 2-3 порядка. Такое их свойство выявляет наличие у них компенсаторной адаптации к недостатку света, которая выражается в адекватных структурно-функциональных изменениях их колоний, полипов и зооксантелл (Dustan, 1979; Zvalinsky et al., 1980; Titlyanov, 1981). Эти изменения направлены на оптимизацию использования кораллом доступной в данном биотопе световой энергии. У разных видов склерактиний такая компенсаторная адаптация выражена в разной степени, что в конечном итоге влияет на их распределение в биотопах рифа с разным уровнем освещенности.
Адаптация кораллов к пониженной освещенности идет по многим направлениям и осуществляется разными механизмами в зависимости от степени дефицита света в местах обитания. При этом в диапазоне ФАРп до 30-10% (освещенность 60-20 Вт/м2) основной механизм адаптации связан с увеличением содержания хлорофилла на единицу площади поверхности колонии. При снижении освещенности в местах обитания ниже 20-10% ФАРп механизм световой адаптации кораллов меняется. Он идет не за счет увеличения концентрации хлорофилла, а главным образом за счет изменения морфологии колоний в направлении повышения удельной численности полипов на ее поверхности и оптимизации перехвата светового потока (Foster, 1979; Jaunert, 1981а; Титлянов, 1987; Лелеткин и др., 1985; Титлянов и др., 1985б).
Основная адаптивная реакция кораллов, обитающих при пониженной освещенности до указанного выше предела 20-10% ФАРп, состоит в повышении в 1,5-3 раза, а иногда и до 7 раз содержания хлорофилла в расчете на площадь поверхности или на вес колонии (Kawaguti, Nakayama, 1973; Falkovsky, Dubinsky, 1981; Titlyanov, 1981; Jaubert, 1981; Dustan, 1982). Такое увеличение плотности фотосинтезирующих пигментов достигается у адаптированных колоний несколькими путями. Прежде всего у адаптированных кораллов возрастает сама численность зооксантелл при общем росте количества живой ткани на единицу поверхности коралла (табл. 12.8) и увеличиваются их средние размеры (Titlyanov et al., 1980). Далее, возрастает концентрация хлорофилла в самих зооксантеллах (Звалинский и др., 1978; Falkovsky, Dubinsky, 1981; Falkovsky et al., 1984; Лелеткин и др., 1985). Рост концентрации хлорофилла в зооксантеллах адаптированных кораллов происходит при этом за счет возрастания плотности упаковки ламелл в их хлоропластах (Машанский и др., 1979; Dubinsky et al., 1984), а также за счет увеличения размеров фотосинтетических единиц (Лелеткин и др., 1980; Титлянов и др., 1981; Falkovsky, Dubinsky, 1981). Способность "теневых" колоний использовать слабый свет связана также с участием в процессе его поглощения дополнительных пигментов типа перидинина, диноксантина и каротина, концентрация которых заметно возрастает в клетках зооксантелл, адаптированных к недостатку света кораллов (Titlyanov et al., 1980; Лелеткин и др., 1988).
За счет увеличения размеров фотосинтетических единиц и более плотной упаковки ламелл зооксантеллы адаптированных колоний кораллов более эффективно используют поглощенную световую энергию, поскольку в этих условиях облегчается транспорт ее к реакционным центрам и возрастает сбалансированность фотосистем (Zvalinsky et al., 1980; Лелеткин и др., 1980) . Эффективность использования солнечной радиации кораллами вообще очень высока. Неадаптированные ветвистые кораллы поглощают до 50% энергии падающей радиации. Кораллы же, адаптированные к недостатку света, могут поглощать до 80% падающей радиации с квантовым выходом фотосинтеза до 16% вместо 1-2% у неадаптированных колоний, что само собой говорит о совершенстве фотосинтетического аппарата их зооксантелл (Falkovsky et al., 1984). Существенно также, что кораллы, адаптированные к недостатку света, характеризуются и более высокой эффективностью использования сильного света выше порога насыщения (Titlyanov, 1981; McCloskey, Muscatine, 1984). Это видно из сравнения световых кривых адаптированных и неадаптированных колоний (см. рис.12.14). В связи с этим у кораллов, живущих в верхних зонах рифа в условиях затенения (в гротах, в нишах) и адаптированных к недостатку света, возрастает эффективность использования проблесков сильного света, попадающего туда от бликов. Таким образом, у колоний кораллов, заселяющих слабо освещенные биотопы рифа, в результате адаптивных изменений появляется возможность с большей эффективностью использовать как слабый, так и насыщающий и сильный свет.
Другой адаптивный механизм, обусловленный жизнью кораллов при недостатке света, связан со снижением у "теневых" колоний интенсивности дыхания в 1,5-3 раза в расчете на массу живых тканей (Davis, 1977, 1980; Титлянов и др., 1985б) и на единицу площади их поверхности (табл. 12.9). Такое снижение позволяет адаптивным формам эффективно расходовать энергетические ресурсы и способствует тому, что величина Р/М у кораллов превышает единицу даже при очень слабой освещенности в местах обитания 10-5 ФАРп (см. табл.12.9, рис.12.17).
Как показали опыты с переносом колоний кораллов из освещенных биотопов в затененные, период адаптации их к недостатку света длится около 2-3 недель (Davis, 1980; Gattuso, 1985). Реакция кораллов на недостаток света выражается также в адаптивных изменениях строения их колоний и в появлении у некоторых из видов специальные структурных приспособлений, которые оптимизируют использование ими слабого света (Barnes, 1973; Graus,Macintyre, 1976; Foster, 1979; Jaubert, 1981а; Lasker, 1981; Fricke, Schuhmacher, 1983; Титлянов, 1987; Титлянов и др., 1985).
У коралла Plerogyra sinuosa, который живет в очень широком диапазоне освещенности, имеются специальные органы, обеспечивающие этому кораллу возможность адаптироваться к разной интенсивности света (Fricke, Vareshi, 1982). На поверхности его крупных полипов имеются фотосинтетические органы типа пузырьков, которые могут меняться в размерах. Ткань этих пузырьков набита зооксантеллами. В случае нужды при низкой освещенности за счет раздувания пузырьков поверхность фотосинтезирующих органов полипов этого коралла может увеличиваться в 5-6 раз. Меняя размеры пузырьков, коралл может менять фотосинтезирующую поверхность адекватной освещенности. Похожие органы фотосинтеза имеют также некоторые симбионтные актинии (Реасае, 1974).