Сильная вариабильность интенсивности питания кораллов in situ была отмечена также у другого вида этого коралла Montastrea annularis (Lasker, 1976). С помощью того же метода экстракции кишечников полипов in situ было установлено, что две морфы этого коралла, обитающие на карибских рифах и имеющие разную величину полипов, имеют разную стратегию охоты и разную способность ловить зоопланктон. Ночная морфа имеет более крупные полипы, чем дневная, и живет на глубинах более 10 м. Полипы этой морфы активны только ночью (см. табл.13.13). Основное время ее интенсивной охоты приходится на время вечерней и предутренней миграции демерсального зоопланктона. Дневная морфа обитает на мелководье, и ее полипы активны круглосуточно. Судя по единовременному наполнению кишечников, они менее эффективные хищники, чем полипы ночной морфы. Однако, поскольку полипы дневной морфы активны крутосуточно, их суммарный суточный рацион оказался близким к таковому полипов ночной морфы, активных только в ночное время.
Расчеты ориентировочных величин отношения суточных рационов к тратам на обмен были выполнены также для коралла Porites по результатам наблюдений за относительным количеством полипов, поймавших жертвы, путем периодического фотографирования его поверхности (Johannes, Терlеу, 1974). У этого коралла с мелкими полипами и короткими щупальцами, являющегося слабым хищником, указанное отношение оказалось близким к 20%. Сходную величину компенсации трат на обмен сообществом кораллов рифов Пуэрто-Рико за счет его хищного питания приводит Глинн (Glynn, 1973). Сравнительное значение автотрофного и гетеротрофного питания кораллов исследовалось в опьпах с кораллом Astrangia danae (Szamant-Froelich, Pilson, 1980; Szamant-Froelich, 1981). Этот коралл обитает в умеренных водах. Часть его колоний в популяциях бывает лишена зооксантелл. Но даже независимо от наличия у них зооксантелл основное их питание — хищное. Автотрофное питание симбионтах колоний этого коралла помогает им переживать периода, бедные зоопланктоном.
В наших исследованиях хищного питания массовых видов склерактиний Б. барьерного рифа Австралии в качестве основного историка меченой пищи использовались науплиусы артемий размером 0,7-0,9 мм, что близко к средним размерам зоопланктона рифа. Однако, как мы уже говорили, кораллы хорошо потребляют и более крупных науплиусов — до 1,4 мм. В то же время науплиусы артемий сильнее большинства планктонных рачков того же размера и легче вырываются на свободу, будучи загарпуненными нематоцистами. Рационы при хищном питании кораллов были определены в опытах продолжительностью 40 мин в ночное время. Ввиду значительной вариабельности величин рационов, получавшихся в разных опытах ввиду индивидуальных различий отдельных колоний, для каждого вида испытуемых кораллов было выполнено 8-10 параллельных определений. С использованием полученных величин
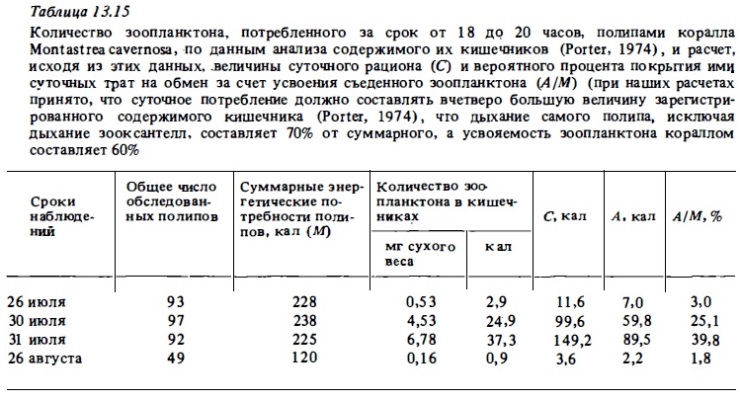
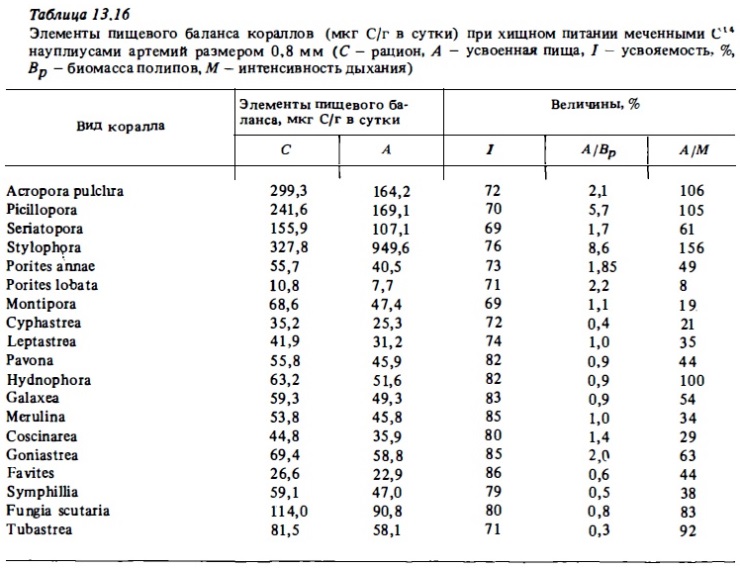
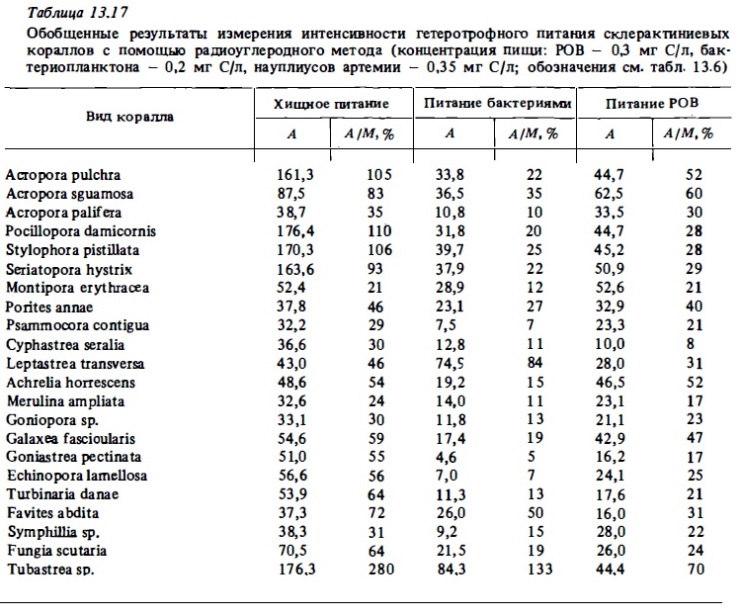
Количество зоопланктона, потребленного за срок от 18 до 20 часов, полипами коралла Montastrea cavernosa, по данным анализа содержимого их кишечников (Porter, 1974), и расчет, исходя из этих данных, .величины суточного рациона (С) и вероятного процента покрытия ими суточных трат на обмен за счет усвоения съеденного зоопланктона (А/М) (при наших расчетах принято, что суточное потребление должно составлять вчетаеро большую величину зарегистрированного содержимого кишечника (Porter, 1974), что дыхание самого полипа, исключая дыхание зооксантелл, составляет 70% от суммарного, а усвояемость зоопланктона кораллом составляет 60% ночных часов рационов, величин усвояемости: животной пищи кораллами (табл. 13.16), а также сведений о суточной ритмике хищного питания кораллов (см. табл. 13.14) были вычислены величины суточного усвоения пищи разными видами кораллов и найдены отношения этих величин к величинам суточного дыхания (табл. 13.17). Результаты опытов доказали способность склерактиний к интенсивному хищному питанию.
За счет хищного питания при концентрациях пищи (науплиусов), близких к верхней границе концентрации зооцианктона в водах над рифами в ночное время, кораллы могли компенсировать от трети до 100% суточных трат на обмен. При этом наибольшие величины суточного усвоения животной пищи в расчете на 1 г веса колонии, а также в процентах от трат на обмен оказались свойственными кораллам с малыми полипами, такими, как Acropora, Pocillopora, Stylophora. У этих кораллов часовой рацион составлял от 6 до 21 мкг С/г и суточное усвоение 134—183 мкг С/г. Будучи рассчитана на биомассу полипов, величина суточного усвоения у этих кораллов составляла 3-5%. Отношение суточного усвоения к суточным тратам на обмен (А/М) часто превышало у них 100%. Это означает, что при концентрации зоопланктона в воде 1-3 г/м3 эти кораллы могут получать за счет хищного питания большую часть необходимой им энергии. Среди кораллов с малыми полипами наименее активными хищниками оказались Montipora и Porites. Слабая активность кораллов из рода Porites, как хищников, отмечалась и другими авторами (Johannes, Tepley, 1974).
Часовые рационы кораллов со средними размерами полипов выражались величинами 2-5 мкг С/г. За счет хищного питания при оптимальной концентрации науплиусов эти кораллы компенсируют 25-50% их трат на обмен (см. табл.13.16, 13.17). Среди кораллов с крупными полипами наиболее активными хищниками оказались кораллы, полипы которых имеют длинные щупальца, такие, как Goniastrea, Turbinaria Echinopora, Favites, Fungia. Их часовые рационы варьировали от 3 до 7 мкг С/г и отношение А/М составляло 55-72%. И все же самым активным хищником среди склерактиний оказался агерматипный коралл Tubastrea sp. Отношение А/М у этого коралла достигало 200-280% при средних величинах часовых ночных рационов 13-18 мкг С/г (см. табл.13.17).
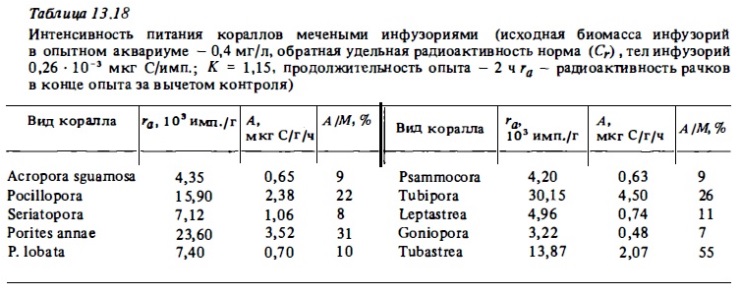
В планктоне вод коралловых рифов иногда в значительном количестве встречаются инфузории, в связи с чем возник вопрос о том, могут ли они потребляться кораллами. Опыты, в которых в качестве пищи использовались меченные радиоуглеродом инфузории, показали, что инфузории, как правило, мало потребляются кораллами. Наиболее активным в этом отношении оказался коралл Tubastrea (табл. 13.18).
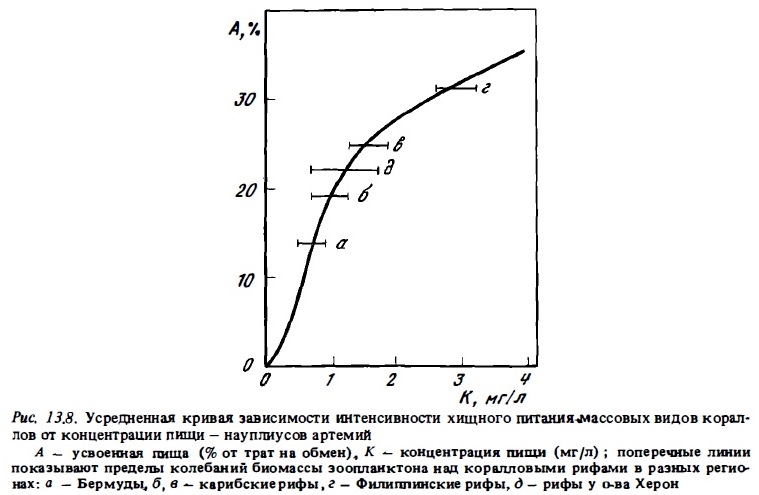
Чтобы решить вопрос о роли хищного питания кораллов в реальных условиях рифа, необходимо было получить данные о зависимости интенсивности их хищного питания от концентрации зоопланктона и измерить реальную ночную биомассу последнего в водах рифа. Результаты этих определений приводятся на рис. 13.7, 13.8. Форма кривых зависимости интенсивности хищного питания кораллов от концентрации зоопланктона мало варьировала у разных их видов. Она также оказалась сходной с аналогичными кривыми при питании кораллов другими видами корма: РОВ и бактериопланктоном (рис. 13.9). Точка насыщения, соответствующая перегибу кривой, достигалась при его концентрации около 1,5 мг/л сырой биомассы. Такая концентрация зоопланктона обычна в воде над рифами в ночное время. На рис. 13.8 представлена усредненная кривая зависимости интенсивности питания кораллов от концентрации зоопланктона. В качестве основной точки отсчета на оси ординат для построения этой кривой была взята средняя интенсивность усвоения кораллами науплиусов артемий при их концентрации 4 мг/л, близкой к верхней границе биомассы ночного зоопланктона в водах рифов. Она была выражена в процентах от суммарных энергетических потребностей кораллов. Последние не равны тратам на обмен, поскольку они включают также траты энергии на рост и на выделение слизи. Величину суммарных энергетических потребностей вычисляли, принимая, что расходная часть баланса энергии у кораллов включает траты 60% энергии на дыхание, 10% на рост и 30% на экскрецию слизи (Sorokin, 1981а; Сорокин и др., 1983). Средняя величина ассимиляции зоопланктона (науплиусов), измеренная нами с помощью радиоуглеродного метода у 27 видов кораллов при оптимальной концентрации корма 3-5 мг/л, составляла около 60% от суточных их трат на дыхание. Выше бышо принято, что дыхание должно составлять 60% от суммарных энергетических потребностей кораллов. Тогда средняя суточная величина усвоения ими зоопланктона при его оптимальной концентрации (3-5 г/м3) должна составлять 36% от их суточных энергетических трат, принятых за 100%.
На построенную таким путем обобщенную кривую зависимости интенсивности питания от концентрации зоопланктона были нанесены значения реальных концентраций ночного зоопланктона, зарегистрированные в водах разных рифов (см. рис.13.9).
реальные концентрации варьируют на разных рифах в среднем от 0,2 до 2 мг/л. При таких величинах концентрации зоопланктона в водах рифов реальная доля хищного питания в энергетическом балансе кораллов должна выражаться величинами от 10 до 30%. На рифах атолла Херон, где проводились описанные исследования питания кораллов, биомасса зоопланктона была близка к 0,5-1 г/м3. В соответствии с кривой (см. рис. 13.8) массовые виды кораллов могли здесь формировать за счет хищною питания 10-20% приходной части своего энергетического баланса.
Способность кораллов к фильтрационно-седиментационному питанию бактериопланктоном была выявлена Ди Сальво в опытах, выполненных с применением изотопов S3 5 (Di Salvo, 1971а, 1973), и нами с помощью С1 4 (Сорокин, 1972, 1973) на биологической станции Гавайского университета в заливе Канеохе на о-ве Оаху. Ди Сальво выдерживал колонии кораллов во взвеси меченых S3 5 бактерий в течение 8-12 ч. Он установил, что при этом кораллы включают в свои ткани до 30-40% внесенной радиоактивности.
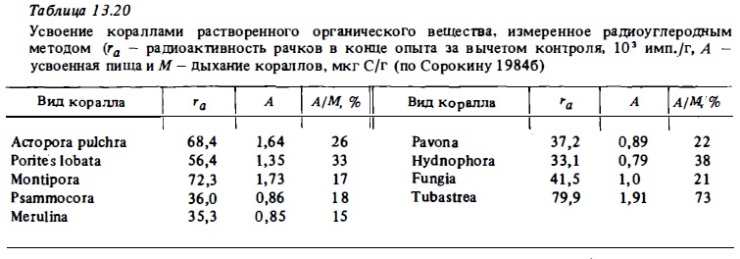
Позже нами были выполнены количественные определения интенсивности потребления и усвоения кораллами бактериопланктона (Сорокин, 1984в). Опыты выполнялись при концентрациях бактериопланктона, близких к верхней границе его концентрации в водах рифа: 1,5-3·106 кл/мл. Результаты некоторых из них представлены в табл. 13.19, а также в табл. 17. В табл. 13.17 приводятся обобщенные результаты опытов по определению интенсивности усвоения кораллами меченого бактериопланктона. Они представлены в ней как в виде абсолютных величин (А), так и в отношениии к тратам на обмен (А/М). Оказалось, что большинство исследованных нами кораллов могут компенсировать за счет питания бактериопланктона 10—20% суточных трат на обмен. У некоторых же видов кораллов интенсивность потребления бактериопланктона была в 2-3 раза выше. К таким видам кораллов относилис: Leptastrea, Hydnophora, Favites.
Для проверки реального усвоения бактерий кораллами мы анализировали также выделение ими меченой углекислоты после кормления их меченным С1 4 бактериопланктоном. Судя по выделению кораллами меченой СО2, органическое вещество потребленного ими бактериопланктона усваивалось полипами, поскольку оно активно включалось в их обмен (Сорокин, 1984в) . Измеренная в ходе тех же опытов зависимость интенсивности питания кораллов бактериями от их концентрации показана на рис. 13.9. Интенсивность питания возрастала почти экспоненциально при росте концентрации бактерий до 0,6-0,8 мг/м3, после чего наблюдался перегиб кривой. Такая концентрация бактериопланктона близка к верхнему ее пределу в водах рифов. Сравнение усредненной кривой зависимости интенсивности питания кораллов от концентрации бактерий (Kb: см. рис. 13.9) с реальными концентрациями бактериопланктона в водах разных рифов позволяет, как и в случае с хищным питанием, оценить относительную долю его в формировании энергетического баланса кораллов. Она оказывается близкой по этим оценкам к 10-15%.
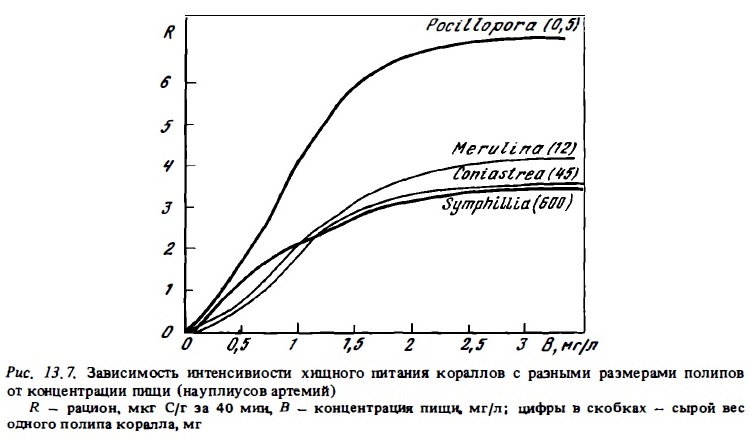
Первые исследователи, установившие факт потребления меченых С1 4 растворенных органических веществ (РОВ) типа глюкозы или глицина кораллами (Stephens, 1962; Goreau et al., 1971), ограничились лишь регистрацией самого факта включения его в состав вещества клеток полипа, не определяя его удельную активность и тем самым не имея возможности оценить скорость его потребления и усвоения. Это делало неясным вопрос о том, может ли усвоение РОВ полипами иметь какое-либо значение в энергетическом бюджете кораллов в естественных условиях. Выполненные нами с помощью радиоуглероциого метода определения интенсивности питания кораллов РОВ выявили достаточно высокую скорость его потребления полипами всех изученных видов (табл. 13.20). При концентрации усвояемого РОВ в воде 1 мг С/л она составляла 20-60 мкг С/г в сутки, что близко к 20-40% от трат на дыхание (см. табл. 13.17). Последние величины получены с использованием обобщенной кривой зависимости интенсивности питания кораллов РОВ от его концентрации в воде (см. рис. 13.9). Концентрация усвояемого РОВ в водах рифа, по данным метода БПК-полного, составляет в среднем 0,2-0,5 мг С/л. В пределах этих концентраций в условиях опыта кораллы за счет усвоения РОВ могли компенсировать до 12-18% своих суммарных энергетических потребностей.
Как мы видели выше, благодаря большому вниманию к вопросам питания и энергетического баланса склерактиний к настоящему времени мы имеем достаточно четкие представления о составе их пищи, интенсивности питания разными видами доступных им кормов, об их усвояемости кораллами. Выяснена зависимость интенсивности питания кораллов от концентрации пищи и на этой основе найдены подходы к решению вопроса о доле разных источников энергии у герматипных склерактиний в. их энергетическом шансе. Так, в условиях станции Херон, где были выполнены анализы зависимости интенсивности питания массовых видов кораллов от концентрации пищи, а также определены естественные концентрации основных ресурсов питания кораллов - зоопланктона, бактериопланктона, РОВ (см. рис.13.8, 13.9), можно уже сейчас вычислить долю разных историков энергии в их энергетическом балансе (Сорокин, 1984в) . Соответствующие расчеты показывают, что в формировании приходной части энергетического баланса склерактиний, заселяющих мелководные участки рифов этого атолла до глубин порядка 10 м, около 60% энергии дает фотосинтез зооксантелл, 20% — хищное питание и 20% — питание РОВ и бактериопланктоном.