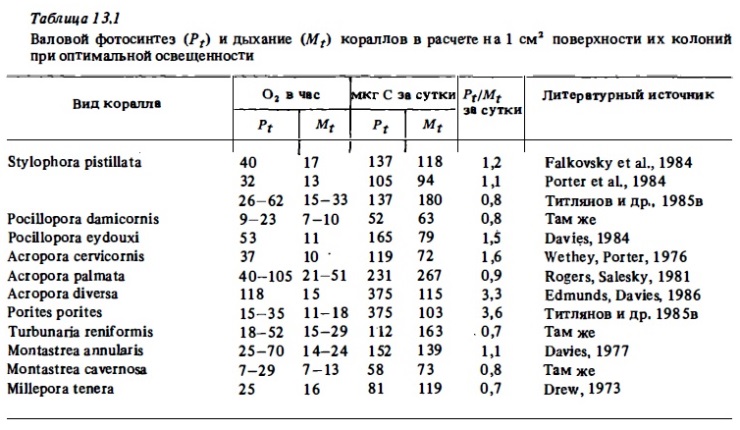
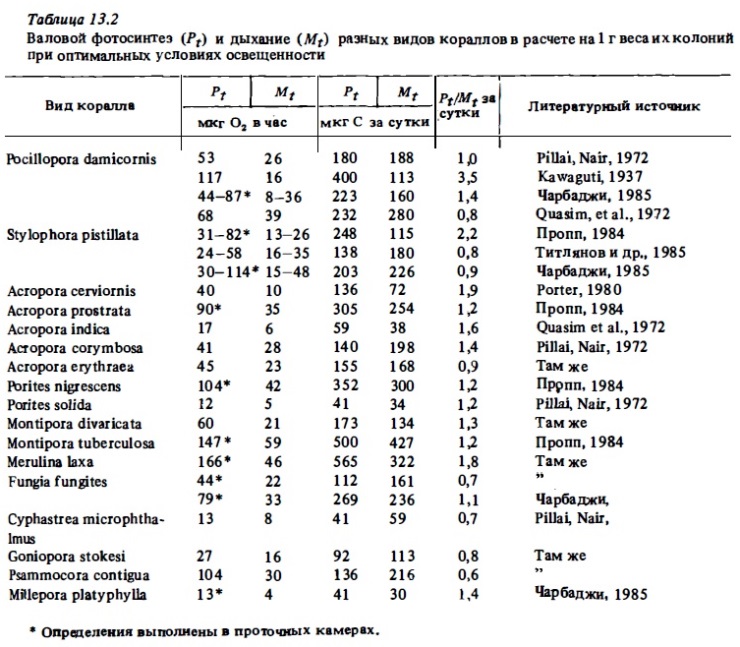
Результаты непосредственных измерений интенсивности фотосинтеза и дыхания кораллов вычислялись разными авторами в расчете на разные интервалы времени (на час, на сутки). Они выражались в единицах кислорода (мг или мл) или углерода в расчете на 1 см2, на 1 м2 поверхности колонии (Davis, 1977; Lasker, 1977), на 1 м2 ее праективной площади (Beyers, 1966), на 1 г ее сырого или сухого веса (Pillai, Nair, 1972; Derw, 1973; Falkovsky, Dubinsky, 1981; Соракин, 1984б), на 1 мг хлорофилла "а", содержащегося в колонии (Wethey, Porter, 1976; Scott, Jitts, 1977; Croaaland, Bames, 1977а; Титлянов и др., 1985а), в расчете на биомассу полипов (Сорокин, 19846), в расчете на 1 мг общего органического углерода, содержащегося в колонии (Lewis, Post, 1982), и, наконец, в расчете на 1 млн клеток зооксантелл (Титлянови др. 1985в). Нетрудно видеть, что такие данные, полученные разными авторами даже для одного и того же вида, трудно сопоставим! между собой. Это касается не только абсолютных величин фотосинтеза и дыхания, но и безразмерных величин отношения Рt: Mt, которые представлены у одних авторав в виде соотношения часовых величин, измеренных в кратковременных 1-6-часовьх опытах в дневное время (Kawaguti, 1937; Kanwisher, Wainwright, 1967; Wells et al., 1973; Мс Closkey et al., 1978), а у других — как соотношение их суточных величин (Wethey, Porter, 1976; Mergner, Swoboda, 1977; Сорокин, 1984б; Титлянов и др., 1985в). Тем не менее, пользуясь об1иирностью имеющегося материала и производя ориентировочные перерасчеты оригинальных данных, можно с достаточной определенностью оценить среднюю интентность фотосинтеза и дыхания массовых видов герматипных склерактиний и многих октокораллов (Lewis, 1977в; 1981а; McCloskey et al., 1978; Muscatine, 1980а).
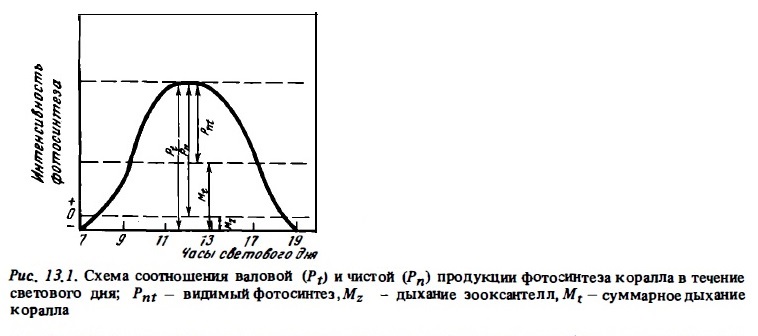
Фотосинтез изолированных зооксантелл и спектры действия их пигментной системы измеряют в их взвеси, помещенной в небольшую освещенную камеру, в которой с помощью электродов регистрируют изменения концентрации кислорода (Лелеткин и др., 1980). Фотосинтез и дыхание кораллов измеряют, помещая колонии или отдельные куски в замкнутые сосуды (метод "склянок") или в проточные камеры. Фотосинтез регистрируют по интенсивности выделения кислорода или по потреблению углекислоты путем измерения колебаний рН или с помощью радиоуглеродного метода, измеряя потребление углерода меченой СО2 кораллом из среды и включение его в состав вещества тканей коралла (Lewis, 1977в; 1981; Мс Closkey, 1978; Muscatine et аl., 1981; Сорокин, 19846; Sorokin, 1986; Титлянов и др., 1985в). Радиоуглеродный метод измерения фотосинтеза дает его величины, которые по оценке одних исследователей близки к валовой (Drew, 1973),а по оценке других — к чистой продукции фотосинтеза (Cooksey, Cooksey, 1972; Taylor, 1973; Scott, Jitts, 1977). Колебания содержания кислорода в опытах по измерению фотосинтеза и дыхания кораллов измеряют методом Винклера или с помощью полярографических электродов. Продолжительность скляночный опытов 1-2 ч. В проточных камерах можно вести более длительные наблюдения. Склянки, а также и проточные камеры экспонирует, как правило, in situ при естественном освещении на рифе. С этой целью рядом авторов были сконструированы автономные агрегаты, которые устанавливались на дне до глубины 50 м. В состав таких агрегатов входят обычно 2 проточные камеры, перистальтические насосы, прокачиваю-щье воду, фотометр, датчики рН, Т°С и кислорода, записывающая аппаратура и блок питания. Эта аппаратура с помощью кислородных электродов, установленных на входе и на выходе потока из камер, регистрирует изменение концентрации кислорода и при прохождении воды через камеру с образцом коралла (Mergner, Svoboda, 1977; Poter, 1980; Chalker et al., 1985). В результате получают кривую суточного хода фотосинтеза и дыхания (в темное время суток), на основании которой вычисляют часовые скорости и суточные величины фотосинтеза и дыхания подопытных кораллов. Пример такой кривой представлен на рис. 13,2. В других случаях проточные камеры устанавливают на борту судна или на берегу, регулируя освещенность нейтральными светофильтрами (Чабарджи, 1985), или камеру устанавливают in situ, а регистрирующую аппаратуру снаружи — на судне или на плоту (Пропп и др., 1983).
В отношении оценки сравнительной адекватности результатов, получаемых в склянках и в проточных камерах, мнения расходятся. Многие сторонники применения проточных камер утверждают, что в склянках, ввиду накопления в них за время опыта продуктов обмена получаемые величины фотосинтеза занижаются до 1,5-2 раз (Dennison, Barnes, 1987; Чабарджи, 1985). С другой стороны, опьпы по сравнению результатов измерений фотосинтеза и дыхания кораллов в автономной проточной камере и в склянках даже при суточной непрерывной экспозиции последних дали сходные результаты (Mergner, Svoboda, 1977). Действительно, в кратковременных 1—2-часовых скляночных опытах трудно ожидать каких-либо серьезных артефактов. Об этом же говорит и общее сходство величин фотосинтеза и дыхания, полученных разными авторами с применением обоих методов (проточного и скляночного) для одних и тех же видов кораллов (табл. 13.1, 13.2).
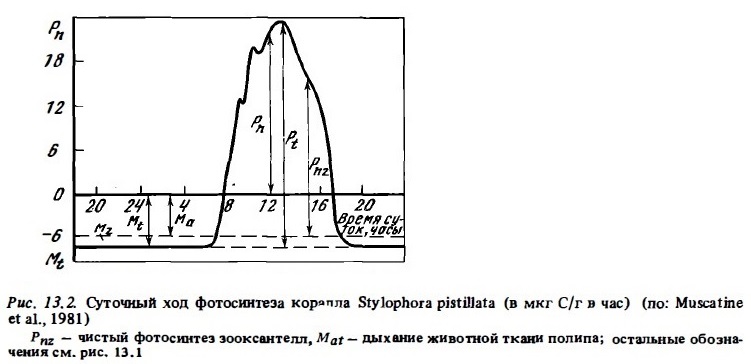
Суть кислородного метода определения фотосинтеза кораллов состоит в регистрации изменений содержания кислорода в воде в присутствии кораллов при экспозиции их на свету и в темноте. При этом прирост кислорода на свету соответствует чистой продукции фотосинтеза Рп, убыль его в темноте — дыханию Mt. Соответственно их сумма дает величину валовой продукции Pt, которая вычисляется в 1 мкг O2 или 1 мкг С в час (см. рис. 13.2). Для перевода величин Pt, получаемых в опытах с O2, в единицы углерода можно пользоваться фотосинтетическим коэффициентом (O2/СO2), равным 1,1. Величины дыхания Mt переводятся в единицы углерода с помощью дыхательного коэффициента (СO2/O2), равного 0,8 (Muscatine et al., 1981), Для вычисления чистой продукции зооксантелл (Pnz) используют соотношение между биомассой зооксантелл и общей -биомассой полипов, принимая, что величины дыхания животных тканей и растительных клеток в составе полипа пропорциональны их биомассе (Muscatine, 1980).
В кратковременных опытах получают, как правило, часовые величин Pt, Рn и Mt. Однако, как мы видели выше, для оценки значения автотрофного питания кораллов в их энергетике, равно как и для оценки их роли в метаболизме донных сообществ, необходимо определить суточные их величины. Поскольку экспозиция суточной продолжительности в замкнутых сосудах практически неосуществима, а в проточных осуществима с большим трудом, суточные величины Pt и Mt получают расчетным путем, измерив часовые их величины в определенный период суток и зная среднее соотношение (К) между часовой продукцией (Pt) и часовым дыханием (Mt) для данного периода суток, во время которого производились измерения (Сорокин, 19846). Величины К, необходимые для расчета фотосинтеза отдельного вида герматипные кораллов или групп кораллов, обитающих в сходных условиях освещенности, мы находили, пользуясь измеренными для некоторые их видов кривыми суточного хода фотосинтеза (см. рис.12.9). Коэффициенты К находили как соотношение площади, ограниченной часовыми интервалами (Si), ко всей площади, ограниченной кривой суточного хода валового фотосинтеза (St). Величина суточиого фотосинтеза (Pt) в этом случае будет равна: Pt = Pth·Ki, где Pth — часовая валовая продукция фотосинтеза и Ki — средний коэффициент К, соответствующий временному интервалу его измерения (см. табл. 12.7). Что касается расчетов суточные величин дыхания Mt исходя из часовых их величин, полученные в кратковременные опытах, то большинство авторов, считая, что ночное дыхание кораллов мало отличается от дневного, вычисляют их прастым умножением часовой величины на 24. Часто в качестве такой часовой величины берут среднюю между ее значениями, полученными в дневных опьпах в темноте и в ночных опытах (Wethey, Porter, 1976; Porter, 1980; Сорокин, 1984б). Правда, существует мнение, что ночное дыхание герматипные кораллов может быть до 1,5—2 раза ниже дневного (Porter et al., 1984; Титлянов и др., 1985в) и что при расчетах суточные величин дыхания необходимо строить кривую его суточного хода. Однако следует учитывать, что разница величин дневного и ночного дыхания может быть артефактом, связанным с тем, что при постановке опытов в условиях рифа обычно используют воду, отбираемую для этой цели в зоне рифов. В дневное время эта вода, как правило, перенасыщена кислородом. Поскольку опыты производятся в затемненных сосудах или аквариумах, избыток кислорода может незаметно теряться в виде пузырьков газа, что может завышать измеряемую по убыли концентрации величину дыхания Mt.
В связи с этим, а также на основании своего опыта мы считаем достаточным для расчетов суточной величины брать часовую его величину, полученную путем усреднения результатов измерений дыхания одной , и той же колонии коралла, выполненные в середине дня и в 9-11 ч вечера.