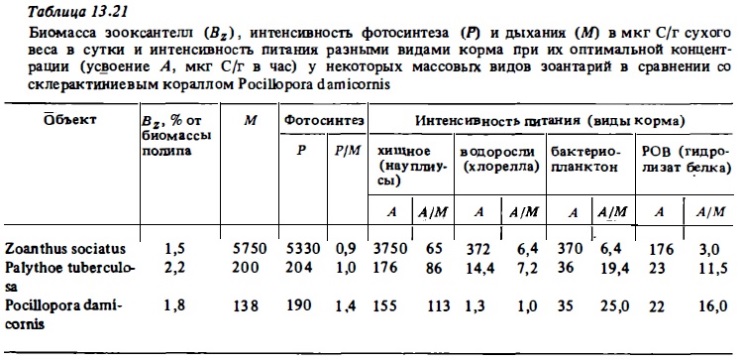
В соответствии с полученными нами данными (табл. 13.21), зоантарии в меньшей степени могут компенсировать траты на обмен за счет фотосинтеза, чем склерактинии. Отношение фотосинтез: дыхание в суточном их балансе у них меньше, чем у последних: 0,9-1 против 1,2-1,5. Следовательно, они должны нуждаться в источниках гетеротрофного питания больше, чем склерактинии. OCia изученных нами вида зоантарий — Palythoe tuberculosa и Zoanthus sociatus способны к хищному питанию. Они усваивают также растительный материал (водоросль хлореллу) и способны потреблять, хотя и менее активно, чем склерактинии, бактериошинктон и растворенное органическое вещество. Триггером пищевой реакции зоантарий, как и у склерактиний, могут быть аминокислоты глицин или пролин (Reimer, 1979а). Антипатарии и цериантарии по своей экологии, включая питание, по-видимому, близки к агерматипным склерактиниям. Основным источником их питания является зоопланктон. Его остатки обнаруживаются в кишечной полости полипов. В условиях эксперимента полипы черных кораллов-антипатарий Antipathes grandis, обитающие на гавайском рифе, ловили, и заглатывали амфипод, копепод, хетогнат (Grigg 1965). Аналогичные наблюдения были выполнены с черными кораллами Cirripathes lutkeni, Antipathes pennacea и А. pedata, взятыми с карибских рифов (Lewis, 1978). Их полипы покрыты ресничным эпителием. Частички пищи они ловят слизевыми сетями, которые затем втягиваются в рот и заглатываются при участии ресничного аппарата орального диска и глотки. Иногда полип использует щупальца для заталкивания пищи в глотку. Полипы этих антипатарий были способны заглатывать очень крупные жертвы. В случае невозможности заглатывания жертвы она опутывается и переваривается вне полипа мезентериальными нитями, выходящими из его рта, как это описано для склерактиний (Lewis, Price, 1976). Ввиду эффективного использования антипатариями механизмов седиментационно-фильтрапионного питания, каковыми являются слизевые сети и ресничный аппарат, следует полагать, что они способны питаться бактериопланктоном и РОВ наряду с зоопланктоном (Lewis, 1978), как это свойственно агерматипным склерактиниями.
Цернантарии по строению полипов сходны с антипатариями и, по-видимому, имеют сходный с ними спектр питания. Судя по наличию в их кишечной полости мощных внеклеточных протеаз, они способны легко переваривать крупные жертвы (Tiffon, Boullion, 1975). Установлено, что они выделяют протеазы и через поверхность эктодермы, обладая, таким образом, возможностью переваривать крупные жертвы, пойманные полипом с помощью нематоцистов, вне его тела (Tiffon, 1975).