Что касается соотношения фотосинтеза и дыхания, то в основном массиве данных преобладают часовые значения их интенсивности и соответственно — часовые отношения Pt/Mt, измеренные для полуденных часов при максимальной интенсивности солнечной радиации. Вполне понятно, однако, что для оценки значения автотрофного питания в энергетике коралла необходимы отношения суточных величин Рt и Мt. В то же время у многих авторов можно встретить выводы об автотрофности питания кораллов лишь на основе найденных ими высоких значений часовых: отношений Pt/Mt, измеренных в середине дня.
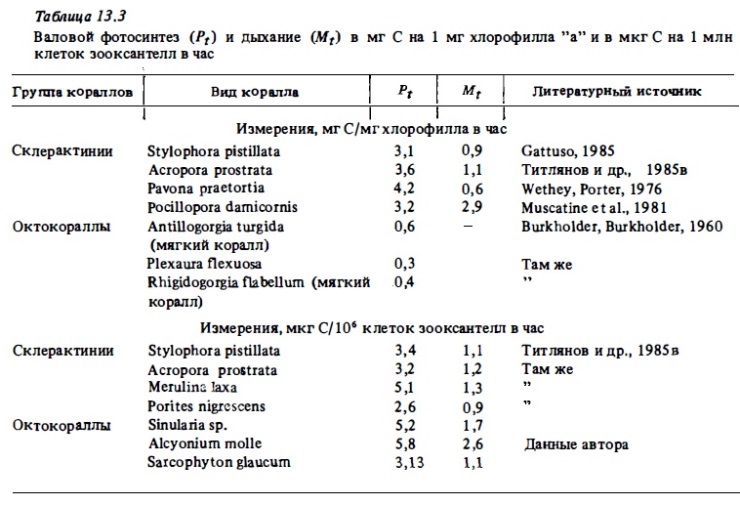
Обобщая данные разных авторов по интенсивности фотосинтеза кораллов, мы постарались показать их в расчете на сутки, исходя из первичных определений, представленных большинством авторов в виде часовых величин. При этом мы использовали полученные нами ранее соотношения суточных и часовых: величин фотосинтеза кораллов (Сорокин, 1984б). В результате такого обобщения нами приводятся таблицы, в которых опубликованные данные суммированы по их размерности: в расчете на 1 см² поверхности коралла (см. табл.13.1), в расчете на 1 г сухого веса колоний кораллов (см. табл.13.2), в расчете на 1 мг хлорофилла и на 1 млн клеток зооксантелл (табл. 13.З). Результаты собственных: измерений продукции фотосинтеза и дыхания склерактиний, мягких кораллов и зоантарий в расчете на 1 г веса колоний и в расчете на 1 мг биомассы углерода полипов представлены в табл. 13.4 и 13.5.
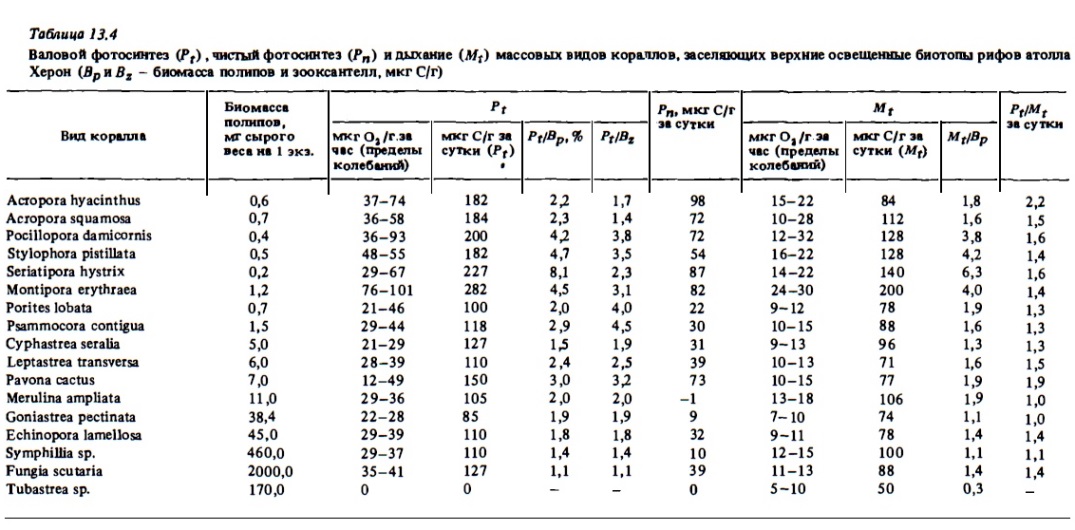
Анализ материала, включая собственные данные, показывает, что в середине дня при оптимальной освещенности в пределах 100-10% ФАРп в расчете на 1 г веса колонии интенсивность валового фотосинтеза большинства склерактиний варьирует в пределах 30-100 мкг О₂ в час. Верхний предел этих велики может достигать 140-160 мкг (Пропп, 1984). Такие величины были зарегистрированы у кораллов Montipora tuberculosa и Merulinaаха в проточных: камерах. Величины валового фотосинтеза ниже 20 мкг О₂/г в час приводятся для некоторых: массивных кораллов, у которых: удельная доля живой биомассы в общей массе кораллов значительно ниже, чем у ветвистых и фолиозных: видов кораллов. В числе таких массивных: форм с низкими значениями валового фотосинтеза на 1 г веса колонии: Cyphastrea, Goniastrea, некоторые виды кораллов Porites и Favites. В наших опытах, выполненных на атолле Херон в единой серии и в сходных условиях освещенности, разброс величин и интенсивности фотосинтеза у разных видов склерактиний был намного меньше, чем это можно встретить в массиве литературных данных (см. табл.13.4 и 13.2). Средние его величины у кораллов с очень разным строением колоний и с разными размерами полипов оказались весьма стабильными и варьировали в довольно узких пределах: 20-40 мкг О₂/г в час у кораллов с более массивными колониями и 30-50 мкг О₂/г в час у ветвистых форм с высокой удельной долей живой биомассы.
В расчете на 1 см поверхности колоний (см. табл. 13.1) часовые величины валового фотосинтеза варьируют в пределах 10-50 мкг О₂, хотя в отдельных случаях были зарегистрированы величины более 100 мкг O₂/г в час (Edmunds, Davis, 1986). Чабарджи (1985) нашел коррелятивную зависимость между величинами фотосинтеза в расчете на 1 г (Pt) и в расчете на 1 см² поверхности с учетом удельной поверхности коралла (S) : Pt = 9,92·1,6·S мкг О₂/г в час. В соответствии с этой формулой величина фотосинтеза у коралла Stylophora pistillata со средней удельной поверхностью около 3 величин Р будет равна (9,92·1,6-3) = 47,6 мкг O₂/г в час, что вполне соответствует величинам Pt, полученным для этого коралла в эксперименте (см. табл. 13.1, 13.2). В расчете на 1 м² поверхности колонии средняя часовая величина валового фотосинтеза герматипных склерактиний близка к 0,2-0,5 г O₂. Интенсивность их валового фотосинтеза в расчете на 1 мин клеток зооксантелл выражается весьма стабильно величинами 10-15 мкг О₂ или 3-5 мкг С в час (см. табл. 13.3). При среднем содержании зооксантедл 2-3 мин/г эти величины и будут соответствовать интенсивности фотосинтеза 20-40 мкг О₂/г в час. Интенсивность фотосинтеза склерактиний в расчете на 1 ми хлорофилла "а" также составляет 10-12 мкг О₂ или 2-4 мкг С в час. Последняя величина соответствует часовому ассимиляционному числу. Судя по этим его величинам, ассимиляционное число зооксантелл in vivo в целом мало отличается от такового у свободно живущего фитопланктона, у которого оно колеблется в пределах 1-10 ми с/мг хлорофилла в час.
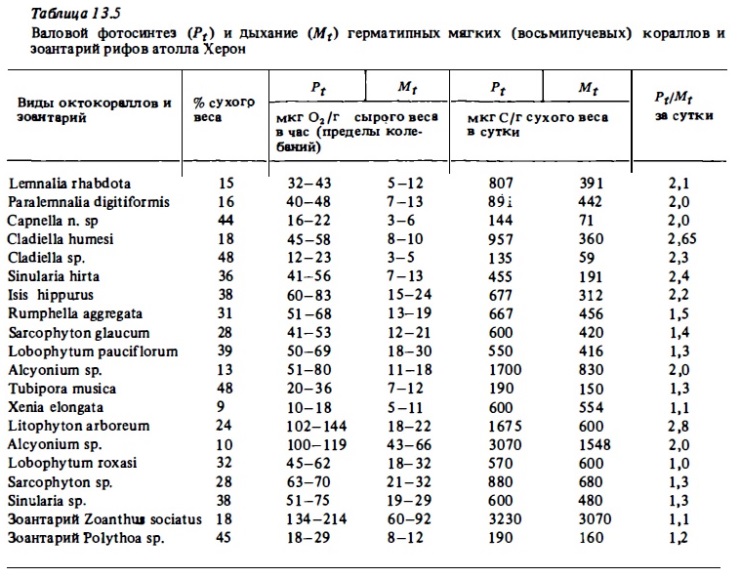
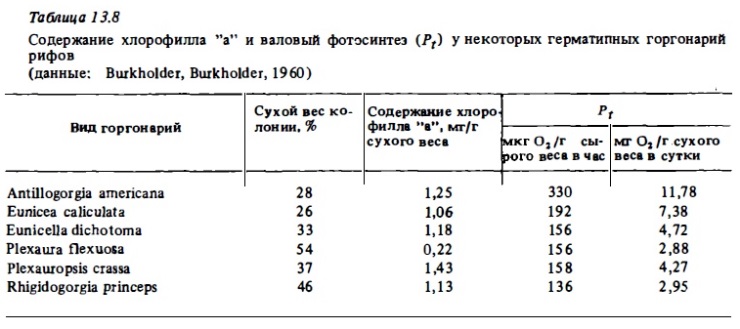
Общий уровень часового фотосинтеза восьмилучевых (мягких) кораллов в соответствии с нашими определениями (см. табл.13.5) в расчете на 1 г сырого веса колоний оказался удивительно сходным с таковым у склерактиний. Его интенсивность выражается у них теми же величинами 30-100 мкг О₂/г в час, что и у ветвистых склерактиний. Причина такого сходства интенсивности фотосинтеза у столь разных по своей морфологии кораллов достаточно очевидна. Она состоит, по-видимому, в том, что зооксантеллы располагаются в полипах слоем толщиною в 1-2 клетки, избегая таким способом самозатенения (Сорокин, 1984а). Плотность распределения полипов в колониях любого коралла такова, что полипы не затеняют друг друга. Поэтому фактически и у склерактиний, и у октокораллов фотосинтетический аппарат достиг в своем развитии оптимальной мощности при данной морфометрии самих колоний. Максимальная интенсивность фотосинтеза среда представителей Anthozoa была зарегистрирована нами у зоантария Zoanthus sociatus, практически лишённого минеральных компонентов скелета. Она достигала у него 214 мкг О₂/г в час или более 2 ми О₂/г сырого веса в сутки (см. табл.13.5).
Суточная величина валового фотосинтеза в среднем в 10 раз выше часовой ее величины (см. табл. 13.1, 13.2). Ее выражают чаще всего в единицах углерода. Величины суточного фотосинтеза, представленные в обобщённых таблицах, были вычислены нами исходя из первичных данных разных авторов о часовом фотосинтезе и использования приведенного выше коэффициента К, равного 10,и величины фотосинтетического коэффициента 1,1. Анализ представленных в них данных, равным образом как и собственных (см. табл. 13.4), показывает, что в расчете на 1 г веса колоний суточная величина валового фотосинтеза ветвистых и фолиозных склерактиний, а также одиноких кораллов функций обычно выражается величинами 100-300 мкг С/г. У некоторых видов (Pocillopora, Merulina) она достигала в опытах 400-500 ми С/г. У массивных. кораллов Goniastrea, Cyphastrea, Porites она близка к 40-80 ми С/г в сутки. В расчете на 1 см² суточные величины фотосинтеза большинства склерактиний варьируют в пределах 50-300 ми C/г (см. табл. 13.1), что составляет 0,5-3 г С/м² (Davies, 1977). Среднее отношение фотосинтезирующей поверхности колоний кораллов к их проективной поверхности близко к 5-10 (Dahl, 1976). Соответственно ожидаемые величины валового фотосинтеза при 100% проективного покрытия дна кораллами могут быть близкими к 3-30 г С/м². Практически же в сообществах кораллов, заселяющих освещенные зоны рифа, они составляют 2-10 г С/м², достигая в отдельных случаях 20-22 г С/м² в сутки. В опытах Баерса максимальные величины валового фотосинтеза двух сравнительно слабо фотосинтезирующих кораллов Porites и Millepora в расчете на проективную поверхность их колоний составили соответственно 3,4 и 10,7 г С/м² (Beyers, 1966). Средние величины суточного фотосинтеза альционарий карибских рифов в расчете на 1 м² проективной площади их колоний составляют 3,7-6,8 г С и у склерактиний тех же рифов 2,7-10,2 С (Kanwisher, Wainwright, 1967). В соответствии с расчетами Тейлора, при 100% покрытия площади дна кораллами на карибских рифах должно продуцироваться за счет фотосинтеза 15 г С/м² в сутки, если экспериментально определенная величина его составляет 260 мкг С/см² в сутки (Taylor, 1973).
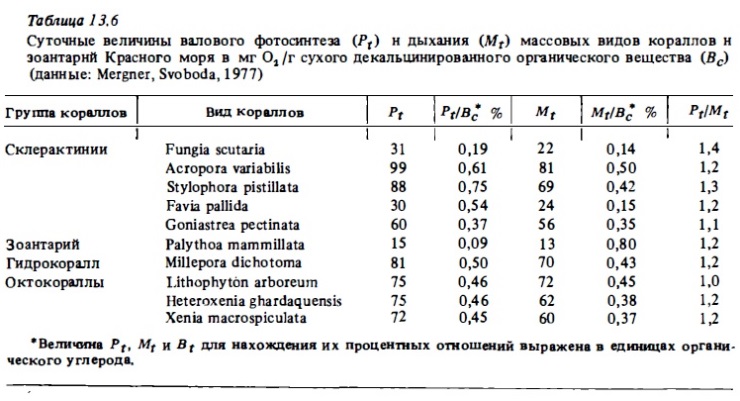
Суточные величины валового фотосинтеза у октокораллов в расчете на 1 г сухого веса их колоний составляют от 0,6 до 3 мг С, что в 2-10 раз выше, чем у склерактиний, хотя в расчете на сырой вес они вполне соизмеримы (табл. 13.6, 13.7). Связано это с большой обводненностью колоний октокораллов, в которых сухой вес составляет 10-40%, а у склерактиний - 70-85%. Наибольшая величина валового фотосинтеза в расчете на сухой вес колоний (4,85 мг С/г в сутки) была зарегистрирована у зоантария Zoanthus sociatus (см. табл. 13.5). В расчете на углерод биомассы полипов (Рt/Вр; см. табл. 13.4) суточный фотосинтез у склерактиний варьировал в пределах 1,1-4,5 мг С и лишь у коралла Seriatopora hystrix с самыми малыми полипами достигал 8 мг C/г. При этом наибольшие величины, как и следовало ожидать, были у кораллов с малыми полипами и соответственно с большей удельной поверхностью.
Чистая продукция фотосинтеза склерактиниевьк кораллов, равная валовой продукции минус дыхание, у большинства из них достаточно велика и составляет при оптимальной освещенности 30-100 мг С/г за сутки или 20-50% от величины их валовой продукции. Чистая продукция фотосинтеза, как и валовая, выше у ветвистых и фолиозных форм кораллов с малыми и средними полипами (см. табл. 13.4). В дневное время в расчете на 1 м² поверхности кораллов часовые величины чистой продукции для многих их видов, обитающих в верхних горизонтах рифов, оказываются достаточно высокими, составляя 150-200 мг С, а у коралла Turbinaria reniformis — до 850 мгС/м² в час (Rogers, Salesky, 1981; Falkovsky et al., 1984).
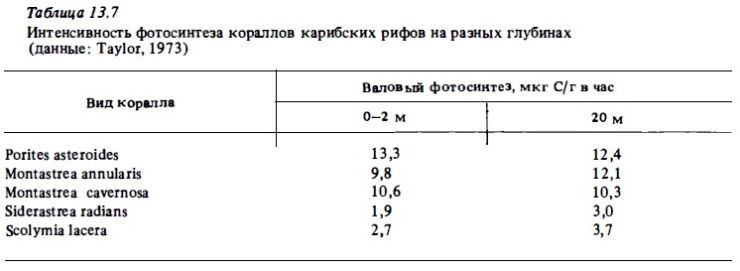
Эффективность использования кораллами солнечной энергии при таком уровне интенсивности их фотосинтеза в освещенных зонах рифа составляет в среднем 0,5-2,5% (Beyers, 1966), возрастая в затененных биотопах до 5-10% (Gattuso, 1985; Muscatine et al., 1985). Как уже отмечалось выше, благодаря механизмам световой адаптации фотосинтез большинства кораллов мало меняется в достаточно сроком диапазоне освещенности, не убывая существенно даже при снижении интенсивности света в местах обитания до порядка величин. В качестве примера в табл. 13.8, 13.8а представлены сравнительные величины интенсивности фотосинтеза кораллов у поверхности и на глубинах до 30 м. Они показывают, что у некоторых видов кораллов фотосинтез более интенсивен при умеренном, чем при полном солнечном освещении.
Интенсивность дыхания кораллов является одним из важнейших эколого-физиологических показателей, который характеризует их энергетические потребности (Davies, 1980; Lewis, Post, 1982). В то же время следует помнить, что величина дыхания как таковая не эквивалентна энергетическим потребностям, а является ix частью, составляющей у кораллов, по разным оценкам, 60-70%. Остальная энергия расходуется кораллом на продукцию' слизи и другой органики, выделяемой в среду, а также на рост и размножение (Muscatine et al., 1984; Сорокин, 1984в; Edmunds, Davies, 1986). Из табл. 13.1-13.6 видно, что у герматипных склерактиний интенсивность дыхания кораллов стабильно выражается вершинами 10-30 мкг О₂/г веса колонии или 7-40 мкг О₂/см их поверхности в час. При этом кораллы с более высоким фотосинтезом имеют, как правило, и более высокий уровень дыхания (McCloskey et al., 1978; Porter et al., 1984). К таким кораллам относятся акропориды (Pocillopora, Stylophora, Seriatopora), а также некоторые фолиозные формы (Montipora, Merulina, Turbinaria). Несомненная причина такой закономерности — более высокая интенсивность транслокации ассимилятов в ткани полипа у подобных форм. Та же причина, по-видимому, обусловливает и отмеченное рядом авторов (Porter et al., 1984; Muscatine et al., 1984; Титлянов и др., 1986в) возрастание дыхания в дневные и вечерние часы, когда зооксантеллы наиболее интенсивно продуцируют и поставляют асиммиляты тканям полипа (McCloskey et al., 1978) . Интенсивность дыхания агерматипных склерактиний, таких, как коралл Tubastrea, вообще лишенных зооксантелл, в 2-3 раза ниже средних величин дыхания герматипных их видов (см. табл. 13.4).
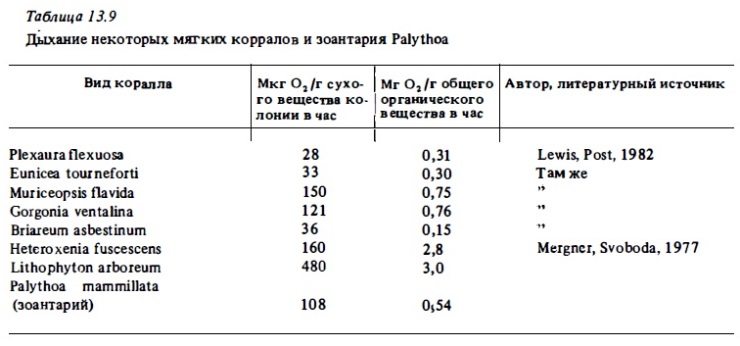
Дыхание мягких кораллов альционарий и горгонарий в расчете на сырой вес колоний в целом выражается величинами, близкими к таковым у ветвистых склерактиний, составляя в среднем 10-40 мкг О₂г/ в час. В отличие 6т склерактиний у октокораллов агермаптные их виды по непонятным причинам характеризуется более интенсивным метаболизмом, чем герматипные, в расчете на 1 г сухого веса колоний. Часовые величины их дыхания варьируют в пределах 30-100 мкг О₂/г сырого веса колоний. В расчете на сухой вес колоний интенсивность дыхания октокораллов выражется величинами 30-500 мкг О₂/г в час или 0,1-3 мг С/г в сутки. Таким образом, диапазон величин суточного дыхания у октокораллов намного выше, чем у склерактиний, ввиду значительных колебаний процента сухого веса их колоний. В то же время в расчете на углерод биомассы полипов интенсивность дыхания октокораллов . варьирует в гораздо меньших пределах. Она составляет 20-50 мг С/г в сутки, что в среднем сходно с аналогичными показателями у ветвистых склератиний. В расчете на 1 г сухого органического вещества интенсивность дыхания октокораллов варьирует в пределах 0,3-3 мг О₂/г в час (табл. 13.9, 13.10). Интенсивность дыхания зооантарий Palythoa и Zoanthus близка к таковой у октокораллов (см. табл. 13.5, 13.6).
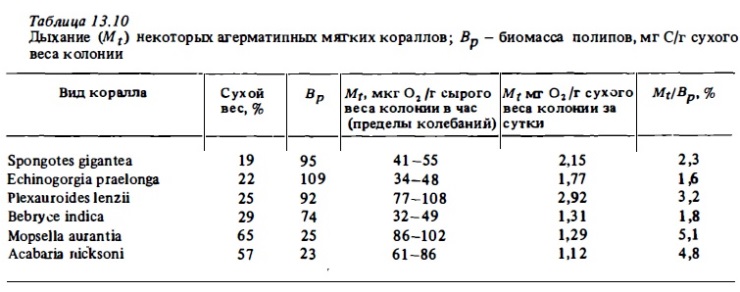
Анализ данных табл. 13.4 показывает, что интенсивность дыхания разных видов склерактиний в расчете на биомассу полипов варьирует в очень узких пределах: от 11 до 42 мг С/г углерода биомассы полипов в сутки (отношение Mt/Bp 0,7- 4,2%). Лишь у коралла с самыми малыми полипами (Seriatopora) отношение Mt/Bp выше 6%. При этом у кораллов со средними и мелкими полипами это отношение в среднем всего в 3 раза выше, чем у кораллов с крупными полипами, масса которых выше на 3 порядка. Таким образом, в колонии коралла интенсивность дыхания фактически мало зависит от индивидуального веса отдельный особей, формирующих колонию. Указанное свойство резко отличает полипы кораллов от свободно живущих беспозвоночный, у который скорость дыхания является обратной степенной функцией индивидуального веса особи. Это важное отличие полипов коралла от свободно живущих животных, несомненно, является следствием их обитания в составе колонии, которая функционирует в значительной степени как единый организм. На основании данных об интенсивности дыхания кораллов в расчете на 1 г общего органического вещества их декальцинированной биомассы некоторые авторы попытались вычислить суточные энергетические их потребности. Но усредненным данным, они оценивают их в 80-200 кал/г С в сутки для склерактиний и в 200-500 кал/г С — для октокораллов (Mergner, Svobcda, 1977; Davies, 1980; Lewis, Post, 1982).
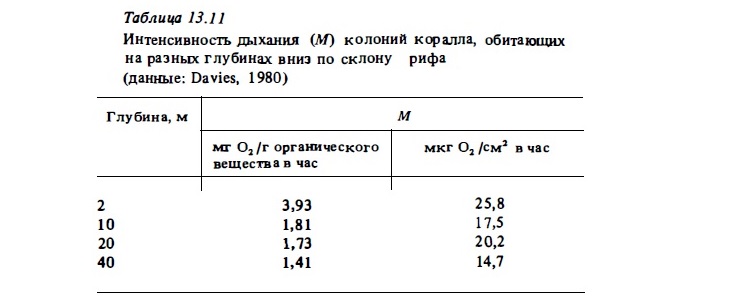
Одна из особенностей физиологии склерактиниевых кораллов, связанная с их адаптацией к пониженной освещенности, состоит в снижении интенсивности дакания по мере убывания величины светового потока в местах обитания (Davies, 1980; Muscatine, Porter, 1977; Лелеткин и др., 1985) (табл. 13.11). Эта реакция наряду с ее явно адаптивным характером может являться отчасти следствием снижения поставки ассимиля-тов зооксантеллами при пониженной освещенности (Muscatine et al., 1984). Адаптивный ее характер подтверждается результатами опытов по трансплантации кораллов из верхних освещеннык горизонтов в глубинные горизонты со слабой освещенностью (Davies, 1980). Опыты показали, что уровень интенсивности дыкания у трансплантированных колоний снижается лишь по истечении 2 недель, после того как коралл пройдет период адаптации к пониженной освещенности.
Как уже отмечалось выше, важнейшим эколого-физиологическим показателем метаболизма кораллов in situ служит отношение валового фотосинтеза к дыханию: Pt/Mt. Это соотношение вычисляют обычно в виде двух его значений: в виде отношения часовых величин максимального дневного фотосинтеза к дневному дыханию и в виде отношения суточных величин Pt и Мt. Часовые величины Pt/Mt варьируют у склерактиний в пределах 2-5. У мягких кораллов и зоантарий они несколько выше: 3-6 (Kan-wisher Wainwright, 1967). Ввиду способности кораллов к световой адаптации это отношение остается выше 1 почти до предельно низких значений освещенности, при которых живет данный вид коралла. Так, у атлантического коралла Synarea convexa отношение Pt/Mt было 1 (1.09) до глубины, где освещенность составляла всего 2,5% ФАРп (Jaubert, 1977).
Суточные величины отношения продукции фотосинтеза к дыханию заметно варьируют в опытах разных авторов даже для одного и того же вида коралла (см. табл. 13.2). Однако, судя по многочисленным результатам их определения разными авторами, для разных кораллов даже при оптимальных условиях освещения они ненамного превышают единицу, а примерно в трети измерений меньше единицы (см. табл. 13.1, 13.2). Мы выползали определения суточных величин отношения Рt/Mt при оптимальном освещении in situ на глубине 1-1,5 м для двух десятков массовых видов кораллов Б. барьерного рифа (см. табл. 13.4). У большинства подопытных кораллов она составляла 1,1-1,4, у двух видов - 1,5, еще у двух - 1,6 и только у одного (Acropora hyacinthus) - 2,2. Весьма существенно, что в отличие от часовых величин отношения Р/М, которые измеряют в середине дня, когда фотосинтез затемненных кораллов максимален, и которые поэтому мало меняются до значительных глубин, суточные величины отношения Pt/Mt, будучи едва больше, а часто и меньше единицы даже при оптимальной освещенности, быстро убывают и становятся значительно меньше в условиях затенения. Основная причина быстрого убывания суточного отношения Pt/Mt с глубиной — сокращение продолжительности светового дня. В качестве примера сравнительного убывания часового и суточного отношений Pf/Mt приведем результаты их определения, выполненного с кораллом Stylophora pistillata. Часовое отношение: на глубине 2 м — 2,6 и на глубине 45 м — 1,0. Суточное отношение: на глубине 2 м — 1,1 и на 45 м — 0,4 (Falkovsky et al., 1984). Итак, в последнем случае энергетические потребности кореша могут удовлетворяться эа счет фотосинтеза менее чем на треть. Остальные две трети потребленной энергии на глубине 45 м коралл должен получать за счет гетеротрофного питания, хотя как мы видели выше, часовое отношение Pf/Mf даже на этой глубине равно у него единице. Поэтому компенсационную точку освещенности для фотосинтеза кораллов следует устанавливать на основании суточного, а не часового отношения Pt/Mt.