а) векторные градиенты параметров среды, такие, как сила волны, течения, уровень прилива, концентрация биогенов, освещенность, содержание взвеси в воде,
б) социальные факторы: группировка особей одного или родственных видов, репродукционные группировки разнополых видов,
в) межорганизменные взаимодействия: комменсализм, агрессия, паразитизм, хищник-жертва,
г) разнообразные стохастические воздействия, включая экстремальные, такие, как тайфуны, наводнения и т .п.
В донных биотопах рифа постоянно существует лимит твердого субстрата ввиду необычайно плотного его заселения сидячей донной фауной, и прежде всего самими кораллами. Так, средний процент покрытия дна в зоне флета составляет 40-60%, достигая на внешнем склоне часто 100% (Ross, Hodson, 1981; Bak Luckhurst, 1980). Даже в лагунах покрытие дна кораллами часто близко к 30-60% (Roy, Smith, 1971). Кораллы при этом обычно занимают 70-90% от всей площади рифа, занятой сидячим бентосом (см. табл.11.5).
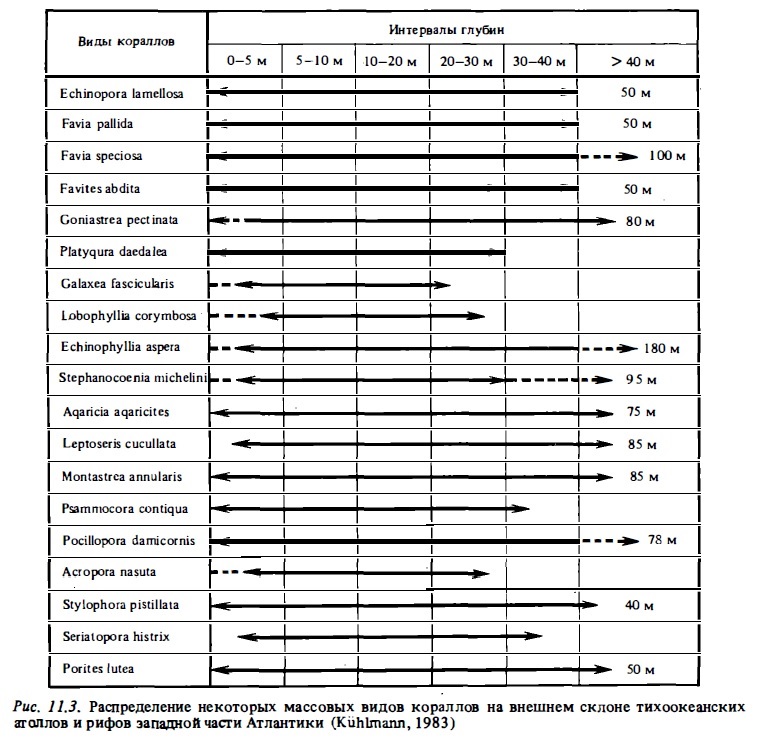
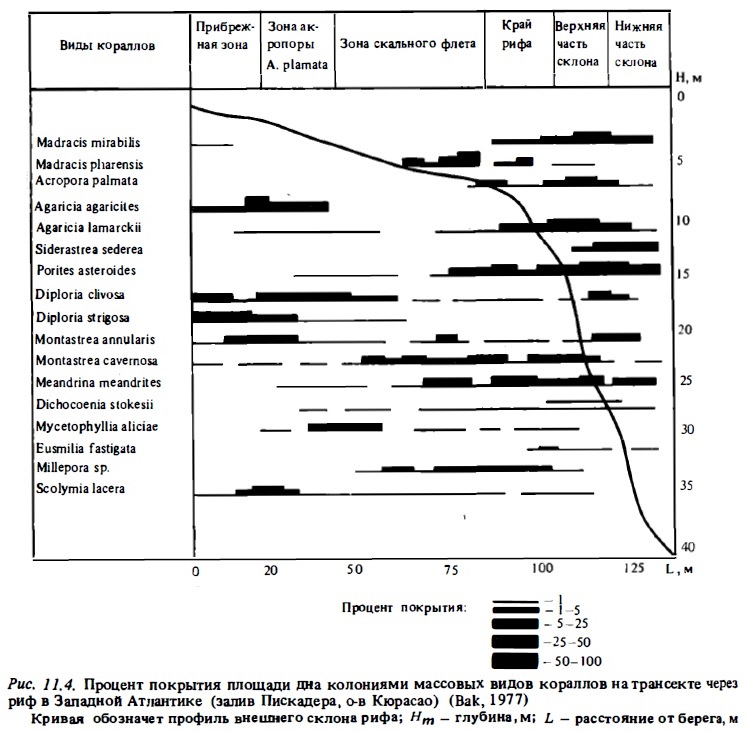
Преобладающая часть кораллов характеризуется широким диапазонам адаптации к воздействию внешних факторов и способна заселять практически: все зоны внешнего склона рифа (рис. 11.3). Об этом говорят кривые распределения числа видов кораллов поперечных разрезах от лагуны и флета до подножия рифа (Kuhlmann, 1983). Пример такой кривой для рассматриваемых рифов приводится на рис.11.4, из которого видно, что на атлантических рифах в пределах глубин от 8 до 60 м видовое разнообразие фауны практически не меняется и лишь на мелководьях сокращается вдвое. На индо-тихоокеанских рифах (рис. 11.5) такое сокращение еще меньше: от 70-90 видов на глубинах 4-50 м и около 60 видов на глубинах 0-4 м. Лишь на приподнятом осы-хающем флете число видов существенно сокращается (Ditlev, 1978). И вся же, несмотря на способность большинства кораллов заселять все зоны рифа, жесткая конкуренция за субстрат приводит к зональному их распределению в биотопах рифа и к появлению доминирующих видов в отдельных зонах. В соответствии с зональным распределением факторов среды от мелководий до подножия внешнего склона рифа меняются и основные параметры, характеризующие структуру их сообществ: видовой состав, состав экоморф отдельных видов, индекс видового разнообразия, проективное покрытие, размер колоний и их возраст (Glynn, 1976; Bradbury, Loya, 1978; Bradbury, 1977; Ditlev, 1978; Sheppard, 1980; Morrisey, 1980; Bradbury, Young, 1981; Sy et al., 1981; Done, 1982).
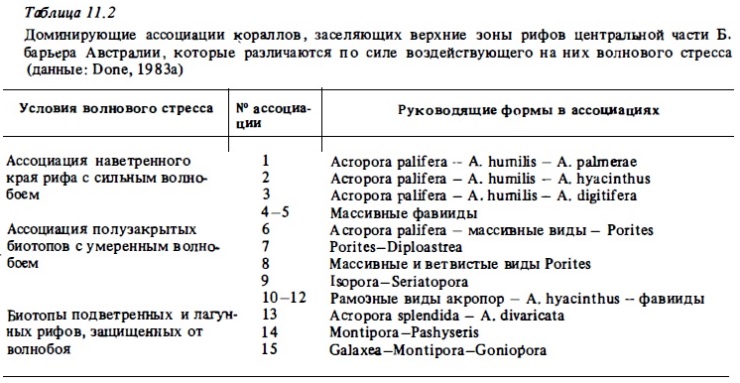
Примеры изменения видового состава кораллов на поперечных разрезах через риф показаны на табл. 11.2. Как видно из этой таблицы, разные зоны рифа заселяют разные видовые группировки кораллов, в составе которых обычно преобладают 2-4 доминирующих вида. Такое доминирование руководящих видов особенно четко выражено в зонах рифах, подверженных наибольшему физическому стрессу. Здесь несколько доминирующих видов могут занимать до половины всей площади дна, покрытой кораллами. При этом многие виды кораллов образуют в разных зонах рифа отличающиеся экоморфы или жизненные формы (Преображенский, 1975; Morton, 1974; Veron, Pichon, 1976; Pichon, 1978; 1981). Некоторые виды кораллов, имеющие в верхних зонах рифа массивные или стол5чатые колонии в нижних зонах склона, в условиях пониженной освещенности образуют идиотские или инкрустирующие (корковидные) колонии. Среди них такие массовые виды атлантических кораллов, как Porites asteroides и Montastrea annularis. В глубинных зонах рифа и на защищенных от волн участках плоские колонии образуют даже ветвистые кораллы, такие, как Millepora platyphilla. Адаптация к разным условиям среды и особенно к условиям освещенности в различных зонах рифа приводит к появлению экоморф, сильно различающихся также и по своим физиологическим характеристикам, например по содержанию в них хлорофилла и по интенсивности обмена (Titlyanov, 1981).
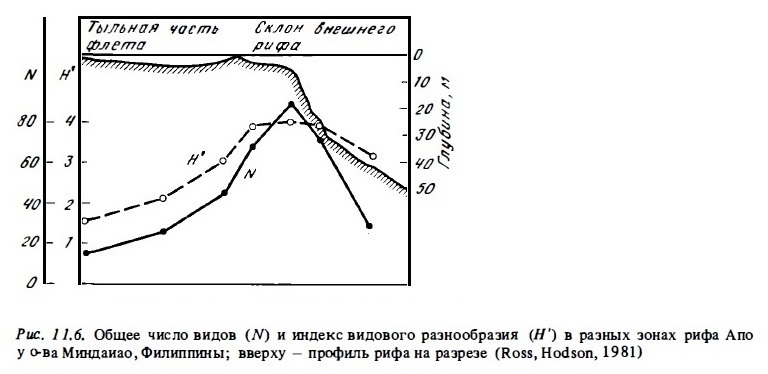
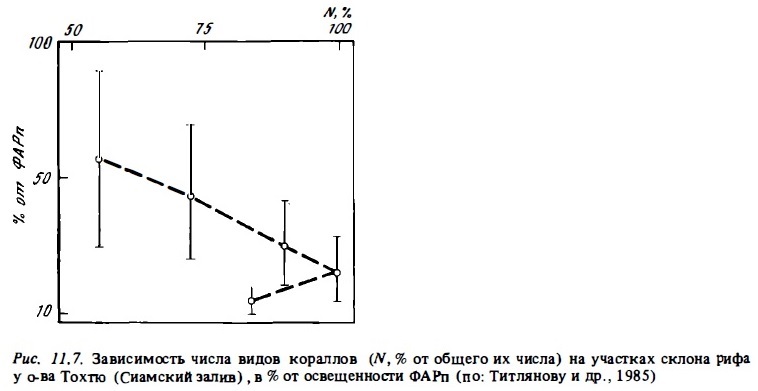
Общее число видов кораллов в составе сообществ и индексы видового разнообразия их сообществ в разных зонах рифа часто заметно различаются (Loya, 1972, 1976). Общее число видов обычно имеет сравнительно низкие значения в мелководных и осушаемых зонах флета и лагуны: 3-5 видов на рифах Атлантики и 10-30 видов на рифах Индо-Тихоокеанского региона. В биотопах наветренного края рифа оно возрастает в 2-3 раза, достигая максимума на внешнем склоне в зоне рва, в зоне опор и в верхней части зоны фор-рифа на глубинах 10-25 м при освещенности 20-10% ФАРп (Porter, 1972; Sheppard, 1980; Ross, Hodson, 1981; Sy et аl., 1981) (рис.11.6, 11.7). Глубже 30 м в нижней части склона число видов кораллов постепенно убывает до 15-40 видов на индо-тихоокеанских рифах и до 5-10 видов — на атлантических. Если наветренная часть флета не подвергается длительной осушке, число видов кораллов на разрезах через такой флет может быть максимальным, достигая на тихоокеанских рифах 100-120 (Wells, 1954) . На флете многих тихоокеанских рифов даже в условиях значительного волнового стресса видовое разнообразие сохраняется на высоком уровне. Примерами таких рифов могут быть наветренные рифы атолла Херон на Б. австралийском барьере (Connel, 1976; Bothwell, 1984) и о-ва Минданао (Ross, Hobson, 1981).

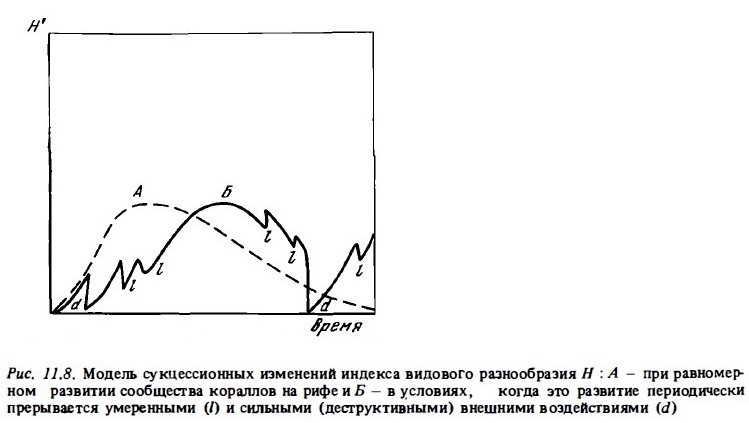
На рис.11.8 представлена теоретическая кривая изменения видового разнообразия сообществ кораллов при равномерной сукцессии в ходе его формирования (кривая А) и гипотетическая кривая Б которая описывает вероятные изменения видового разнообразия в ходе сукцессии, прерываемой умеренными (е), сильными (m) и катастрофическими внешними воздействиями (d). Гипотеза нарушений, объясняющая высокое видовое разнообразие кораллов в биотопах рифа, разделяется в настоящее время большинством специалистов (Connell, 1978, 1983; Grigg, 1983). Тем не менее следует подчеркнуть, что эта гипотеза, по-видимому, приложима в большей степени к сообществам верхних горизонтов рифа, которые сильнее испытывают периодическое воздействие катастрофического стресса. В нижних горизонтах рифа причину высокого видового разнообразия кораллов можно видеть скорость в высоком уровне видовой трофической специализации, как это предсказывает теория (Slobodkin, 1968). Таким образом, в разных зонах рифа и разных рифах в зависимости от местных условий видового разнообразия кораллов обеспечивается под влиянием одного или другого из описанных выше механизмов (Harriott, 1983).