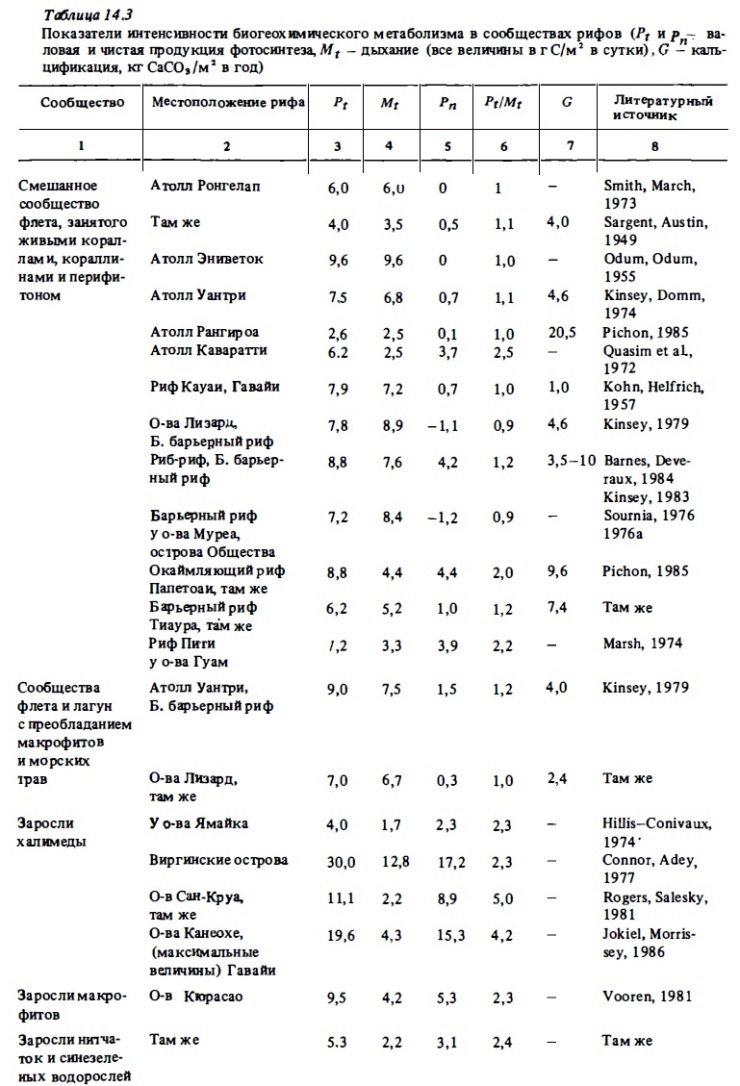
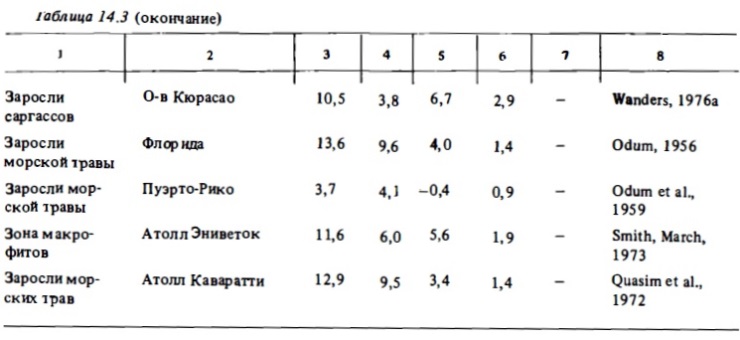
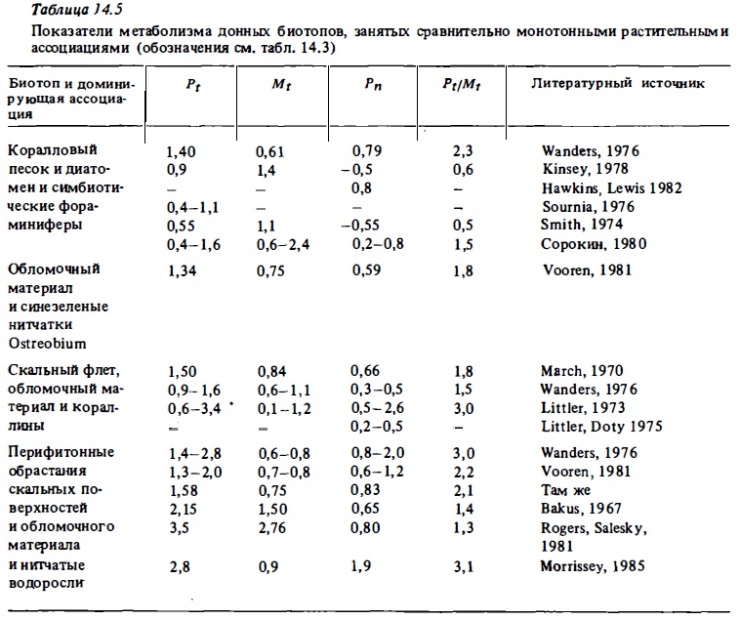
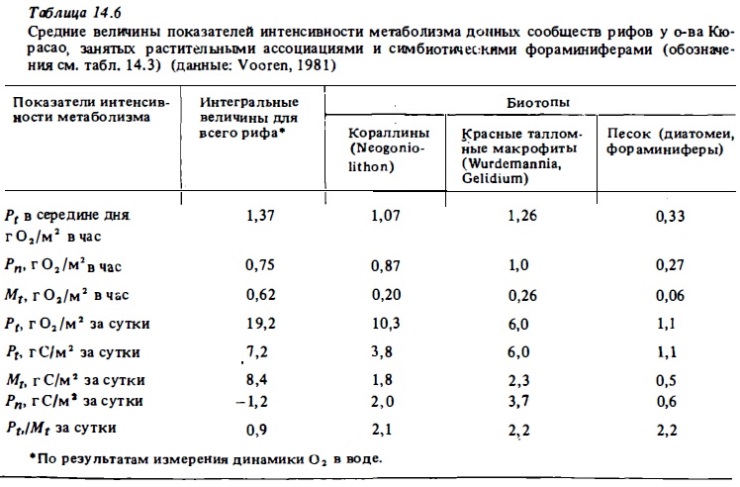
Средний уровень фотосинтетической продукции (4-9 г С/м²) характерен для смешанных донных сообществ риф-флета, занятых живыми кораллами, макрофитами, нипатками, кораллинами (табл. 14.3-14.6). В сообществах внешнего склона валовой фотосинтез варьирует в пределах 0,6-7 г С/м² в зависимости от глубины и уровня освещенности (Davis, 1977). Согласно этому автору, величина валового фотосинтеза, выражающаяся в верхах освещениых горизонтах значением 3 г С/м² в сутки, на глубине 40 м снижается до 0,6 г С/м² при относительно небольшом сражении интенсивности дыхания. Вполне очевидно, что при этом в сообществе ниже горизонтов внешнего склона начинают доминировать гетеротрофные процессы. Валовой фотосинтез участков флета, внешнего края рифа и склона, занятых кораллинами, как правило, относительно невысок: 0,5-З,5 г С/м² в сутки (см. табл. 14.3). В донным биотопах лагун атоллов и барьерным рифов, занятым живыми кораллами, макрофитами, известковыми водорослями (халимедой) и морскими травами, валовой фотосинтез достаточно высок и выражается величинами 5—15 г С/м² в сутки. В биотопах лагуны, занятым песком или обломочным материалом, продукция фотосинтеза составляет 0,4-4 г С/м² в сутки (табл. 14.7, см. также табл. 14.3-14.6). Наши опыты показали, что глубина слоя фотосинтеза в коралловым песках близка к 2-3 см. Основными первопродуцентами в них являются бентосные диатомеи, синезеленые водоросли, нитчатки и симбионтные форамтциферы. Коралловые пески характеризуются высоким содержанием хлорофилла "а" (100-900 мг/м²). Часовое ассимиляционное число в верхнем слое песка может варьировать в пределах от 0,2 до 1,5 мг О₂ на 1 мг хлорофилла в час (Sournia, 1976). Хотя уровень продукции фотосинтеза в таких биотопах в 2-3 раза ниже, чем в сообществах флета и патч-рифов, их вклад в суммарную автотрофную продукцию экосистемы рифа может быть достаточно велик. Например, в экосистемах атоллов благодаря относительно большой площади их лагун (60-90%) вклад сообществ лагун в суммарную первичную продукцию составляет 20-50% (см. табл. 14.7). На одном из микро-атоллов в лагуне атониа Уантри вклад перифитонных обрастаний обломочного материала, песка и мертвых кораллов в суммарную первичную продукцию достигал 80 (Borowitzka et al., 1983).
Наряду с оценками величин валовой (Рt) и чистой (Рn) продукции фотосинтеза в сообществах рифа определенный интерес представляет оценка их первичной продукции (Рnp). В данном случае под первичной продукцией подразумевается чистая продукция самих растений, а не всего сообщества в целом. Между величинами Рn и Рnp в отношении сообществ рифа существует аналогия с величинами Рn (чистая продукция колонии) и Pnz (чистая продукция зооксантелл) в отношении кораллов. Величина первичной продукции сообщества рифа, включающего, как и колония кореша, автотрофные и гетеротрофные компоненты, будет равна разности между валовой продукцией фотосинтеза сообщества и дыханием его растительных компонентов (Мр):Рnp = Рt-Мр. Аналогичньм образом Pnz кораниа равна валовому фотосинтезу (Рt) минус дыхание зооксантени. Причем в последнем случае дня величины Pnz необходимую величину Mz находят расчетным методом. Сходным путем вычисляют и первичную продукцию сообщества рифа. Необходимую дня расчетов величину Мр находят, принимая, что она составляет 15-20% от величины Р:Рnp = 0,85Pf (Atkinson, Grigg, 1984).
Вычисляемая таким путем первичная продукция сообщества рифа имеет важный экологический смысл. Она соответствует величине автотрофной продукции, которую получают в свое распоряжение гетеротрофные компоненты этого сообщества. Часть первичной продукции они используют для своего метаболизма (Мh). Неиспользованный остаток первичной продукции и будет соответствовать чистой продукции фотосинтеза сообщества Рn:Рn = Pt-Мр-Mh, поскольку Мр+Mh=Mt. Поэтому именно величина Рnp (первичная продукция)) соответствует вкладу автотрофной продукции в общий энергетический вход экосистемы. В связи с этим при сравнении уровня продуктивности разных экосистем следует использовать годовые величины их первичной продукции в той трактовке, как она была дана выше, т.е. как чистую продукцию самих растений-первопродуцентов.
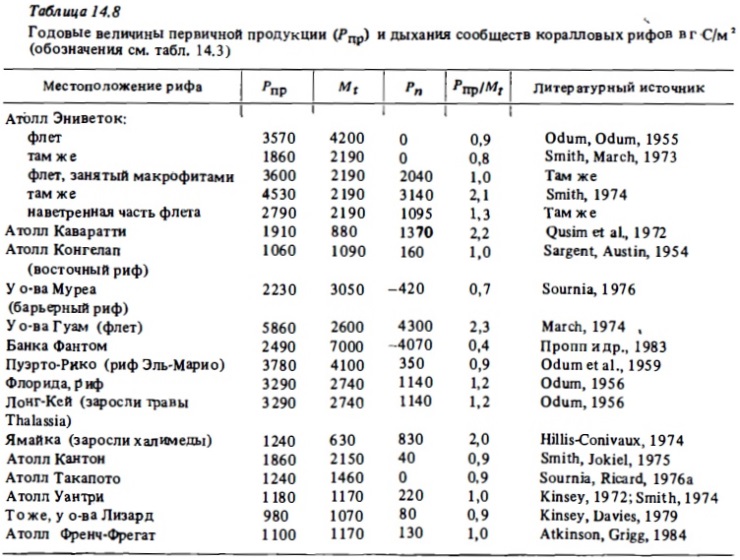
В табл. 14.8 представлены вычисленные, как описано выше, годовые величины первичной продукции фотосинтеза сообществ коралловых рифов, а также величины чистой продукции и отношение Pt/Mt. Как видно из представленных данных, первичная продукция сообществ рифов близка к рекордным ее величинам, зарегистрированным в природных растительных сообществах: тропических лесах, манграх, зарослях бурых водорослей, на шельфе умеренных вод (келпа) (Lewis, 1977а, 1981а). В сообществах флета, занятых корешами и макрофитами, она составляет в среднем 1,5-3 кг Соргна 1 м², что эквивалентно 3,5-7 кг сухого или 20-40 кг/м² сырой биомассы. В отдельных случаях годовая валовая продукция фотосинтеза в биотопах рифа достигала 4—6 кг С/м² или около 80 кг сырой биомассы на 1 м². Эти величины почти на порядок выше годовой первичной продукции экосистем пелагиали в самом продуктивном районе океана — в районе квазипеременного апвеллинга, где она выражается величинами 500-900 г С/м² в год. Лишь во время отдельных вспышек "цветений" фитопланктона первичная продукция может достигать там 5-8 г С/м² в сутки, но такие периоды относительно кратковременны. В то же время в сообществах рифов высокий ее уровень сохраняется относительно постоянно в течение сезона, испытывая значительные колебания лишь на высокоширотных рифах, где имеют место ощутимые снижения температуры воды в зимний период (Kinsey, 1985).
Средние величины годовой первичной продукции, вычисленные дня целых рифовых систем, как правило, оказываются в 1,5-2 раза ниже оценок ее уровня, которые были получены обобщением основного массива данных, относящихся преимущественно к высокопродуктивным биотопам риф-флета и патч-рифов. В соответствии с этими данными средняя первичная продукция этих биотопов составляет около 2,3 кг С/м² в год при уровне чистой продукции сообществ (Рn) 0-0,3 кг С/м² и отношении Pt/Mt (Kinsey, 1983) (см. табл. 14.7-14.8). В большинстве рифовых систем значительная доля их общей площади занята лагунами, в которых первичная продукция фотосинтеза в среднем в 2-4 раза ниже, чем в упомянутых ранее зонах рифа, занятых твердым субстратом. Поэтому следует согласиться с мнением Кинзи, что средняя годовая первичная продукция рифовых систем должна была бы выражаться величинами 1,1—1,4 кг С/м², что соответствует результатам прямых ее определений на ряде атоллов. Так, наиболее репрезентативные полносезонные определения валовой, первичной и чистой продукции фотосинтеза рифовой системы атолла Френч-Фрегат дали следующие их величины: 1,3; 1,1 и 0,13 кг С/м² (Atkinson, Grigg, 1984). Однако при выборе средней величины годовой первичной продукции рифовых систем, необходимой для оценки глобальной продукции коралловых рифов, следует учитывать, что упоминавшиеся данные об ни продуктивности были получены главным образом для атоллов, где действительно лагуны занимают большую часть площади всей системы. В то же время основная масса рифов представлена барьерными и окаймляющими рифами, где площадь лагун не столь велика и где сообщества самих лагун продуктивнее лагун атоллов благодаря более интенсивному развитию в них кораллов и водной растительности. Эти соображения заставляют нас оценить среднюю величину годовой продукции фотосинтеза коралловых рифов в 1,8 кг С/м² и величину их первичной продукции в 1,5 кг С/м².
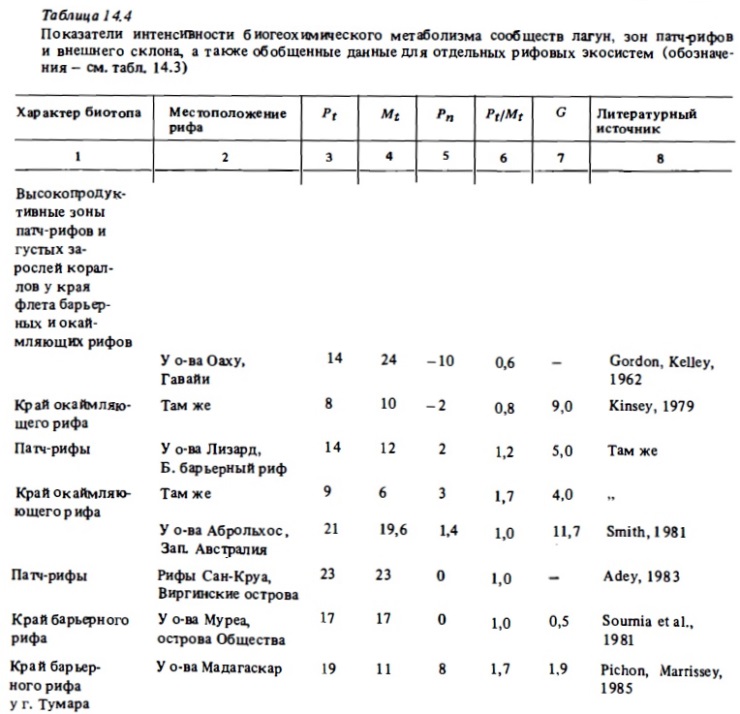
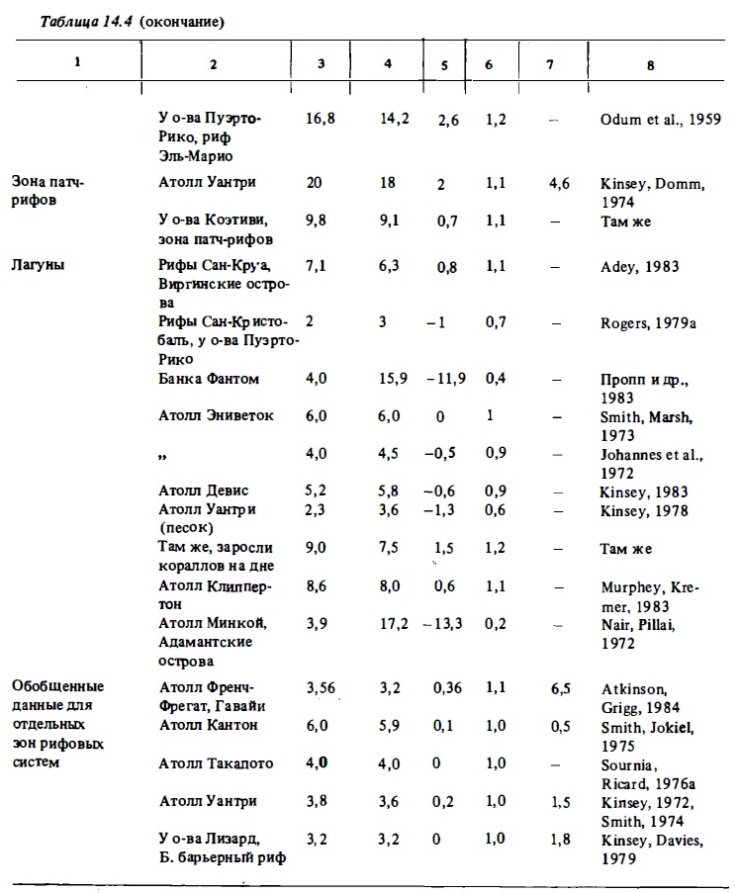
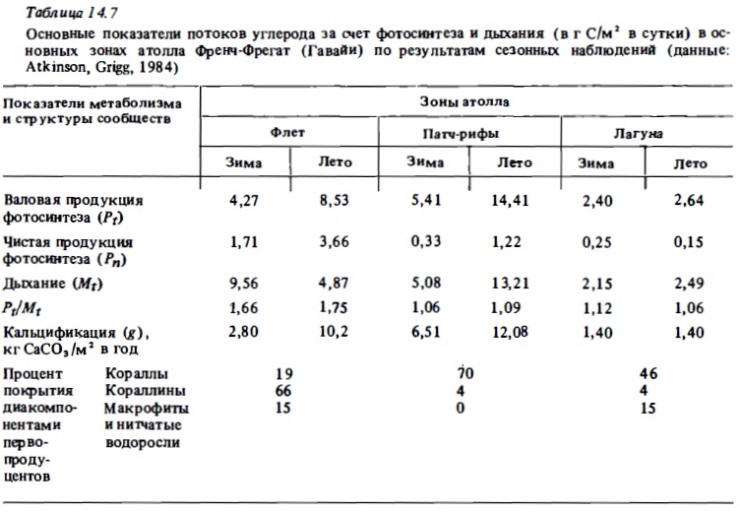
Изучение сезонных изменений метаболизма разных зон рифа бьшо выполнено на атоллах Френч-Фрегат (Atkinson, Grigg, 1984), на атолле Уантри (Kinsey, 1978, 1979, 1985), на рифах Аброльхос (Smith, 1981). Результаты, полученные первыми из перечисленньк авторов, приводятся в табл. 14.7. Максимальная валовая продукция в летний период (14,41 г С/м² в сутки) наблюдалась, как и следовало ожидать, в зоне патч-рифов с доминированием кораллов. Достаточно высокой она было на флете со смешанным сообществом (8,53 г С/м²) и самой низкой - в лагуне, занятой песком с редкими кораллами и макрофитами (2,64 мг С/м²). Атолл Френч-Фрегат (Гавайи) расположен у границы северного тропика. Температура воды в этом районе меняется от 27 С летом до 23°C зимой. Исследоваш1я выявили здесь значительные сезонные колебания интенсивности метаболизма рифа. В зонах флета и патч-рифов зимой продукция фотосинтеза снижалась по сравнению с летним периодом в 2-2,8 раза. В той же пропорции в этих биотопах снижается интенсивность дыхания, интенсивность же кальцификации на флете снижается зимой более чем в 3 раза.
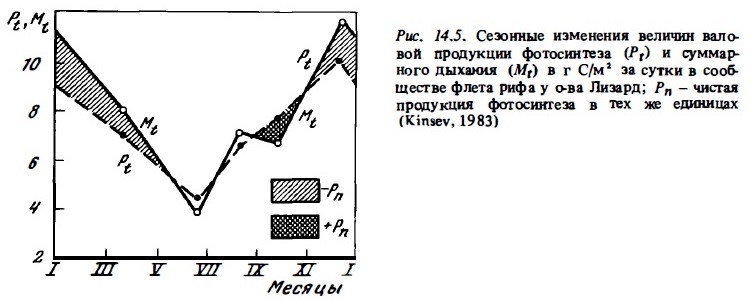
Сравнительные данные об интенсивности фотосинтеза, дыхания и кальцификации в сообществах некоторых других северных и южных рифов с выраженными сезонными изменениями температуры (рис. 14.5) также демонстрируют существенные сезонные колебания величин перечисленных показателей интенсивности метаболизма. Согласно результатам наблюдений, выполненных на рифах Сан-Круа, расположенных на 18° с.ш., зимой фотосинтез и дыхание сообществ флета снижались в 1,8 раза (Adey, Steneck, 1985; цит. по: Kinsey, 1985). В свете этих данных при растете годовых величин продукции фотосинтеза, дыхания я и кальцификации для северных и южных рифовых систем безусловно необходимо учитывать вероятность сезонных их изменений в пределах 1,5—3 раза. В то же время большая часть годовых величин их первичной продукции и деструкции органического вещества, которые приводятся в литературе (см. табл. 14.8), получен умножением суточных величин, определенных в кратковременных наблюдениях, на число дней в году. Отношение же валового фотосинтеза к дыханию (Рt/Мt), как оказалось, не испытывает существенных сезонных колебаний (см. табл. 14.7, рис. 14.5). Лишь на атолле Уантри, расположенном у границы южного тропика, происходило снижение отношения Рt/Мt от 1,0 зимой до 0,8 летом в связи с ростом величины Мt в прогретой воде. Таким образом, трофический статус отдельных биотопов не подвержен существенных сезонным изменениям.
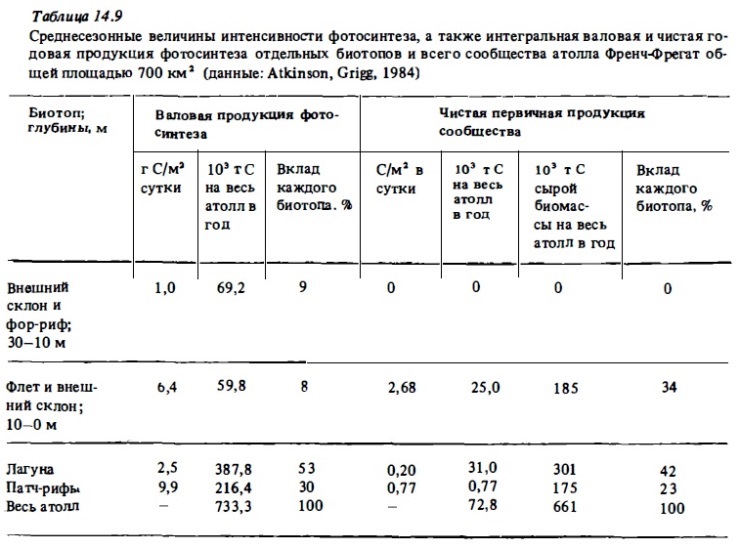
Все донные биотопы рифа характеризуются необычайно высоким уровнем гетеротрофного метаболизма. Общая интенсивность деструкции органического вещества составляет 3-8 г С/м². В отдельных зонах рифа она может достигать 15-18 г С/м² в сутки, что соответствует суточной деструкции 40-50 г/м² сухого органического вещества или 600-700 г/м² сырой биомассы. Благодаря столь высокой деструкции отношение Рt к Mt в большинстве случаев бывает близким к балансу, выражаясь чаще всего величинами 0,8-1,5. Особенно четко эта сбалансированность процессов проявлялась при интегральных определениях валового фотосинтеза и дыхания целых рифовых систем (атоллов), которые включают биотопы с преобладанием автотрофных или гетеротрофных процессов (табл. 14.9). Как видно из табл. 14.3-14.6, отношение Рt/Mt, близко к единице (1-1,5) в сообществах флета и патч-рифов, в приглубык участках внешнего края рифа и верхней части внешнего склона рифа, где преобладают живые кораллы. Соответственно чистая продукция фотосинтеза в таких зонах, как правило, бьта относительно невелика и не превышала обычно 0,5-2 г С/м² при валовой продукции 5-20 г С/м² в сутки (Kinsey, 1978, 1979, 1985). При зтом в подобньк биотопах атоллов отношение Рt к Mt и чистая продукция фотосиитеза бьти, как правило, выше, чем в биотопах барьерный рифов у высоких островов, где это отношение часто меньше единицы, а чистая продукция имеет знак минус (Жариков, 1987) (см. также табл. 14.3, 14.4). Снижение отношения Рt к Mt в последнем случае указывает на поступление внешнего органического вещества с терригениыш стоком с островов.
В зонах флета, где преобладают макрофиты или морские травы, валовый фотосинтез, как правило, в 3-5 раз превышает ныхание, а величина чистой продукции достигает 3-5 г С/м² в сутки. В зонах рифов с преобладанием мяских грунтов (дно каналов, лагун) дыхание во многих случаях превышает фотосинтез и отношение Рt/Mt бывает меньше единицы Источником органического вещества дня избыточного дыхания в данном случае является его поступление из зон флета и патч-рифов, а также с участков дна, занятых манграми, морскими травами и макрофитами, где производится избьпок чистой авто-трофной продукции (Сорокин, 1973в; Kinsey, 1985, 1985а; Жариков, 1987). Это органическое вещество поступает из зон высокой первичной продукции преимущественно в ввде растительного детрита, а также слизи кираллов, оформленной как органические агрегаты и фекалии рыб. Такой перенос органического вещества из "автотрофных" биотопов фаций рифа в "гетеротрофные" обеспечивает энергетическую взаимосвязь, объединяющую биотопы рифа в единую экосистему.
Существенное значение для оценок энергетического баланса и соотношения разнонаправленных процессов биогеохимического метаболизма в сообществах рифов имели уже упомииавшиеся выше сезонные исследования энергетики экосистемы атолла Френч-Фрегат из группы Гавайских островов, которые выполнялись в рамках проекта "ЕСОРАТН" (Polovina, 1984; Atkinson, Grigg, 1984). В ходе этих исследований были определены величины валовой первичной и чистой продукции фотосинтеза, дыхания и кальцификации в четырех основных биотопах атолла: на флете, в зоне патч-рифов, в лагуне и в зоне внешнего склона в разные сезоны года. Определив площадь, занимаемую перечисленными выше биотопами, исходя из найденных величин продукции, были вычислены суммарные величины валовой и чистой продукции донных сообществ на всей площади атолла, а также вклад каждого из этих биотопов в суммарную продукцию (см. табл. 14.9). Валовая продукция фотосинтеза в расчете на всю площадь атолла (700 км²) составляет более 700 тыс. т углерода, чистая автотрофная продукция сообщества составила почти 70 тыс. Что касается относительного вклада биотопов в суммарную,валовую и чистую продукцию, то наибольшим он был у биотопов лагуны, которые характеризуются самой низкой продукцией, но занимают более половины общей илощади атолла. Наименьшим этот вклад был у сообщества внешнего склона и флста. Чистая же продукция сообществ флета была близка к нулю ввиду высокого уровня дыхания.
Биогеохимический метаболизм углерода включает наряду с фотосинтезом, дыханием и ростом живых тканей еще и процесс роста известкового скелета донньгх животных и известковых водорослей — кальцификацию (Smith et al., 1978). Ее интенсивность в сообществах рифа велика и варьирует в пределах 1-20 кг СаСO₃ на 1 м² площади дна в год. Это соответствует суточной кальцификации 3-55 г СаСO₃ или 0,2-4 г С/м² в сутки, что в среднем близко к нижним значениям валовой, но соизмеримо с чистой продукцией фотосинтеза.
Протекающие в сообществах коралловых рифов процессы продукции и деструкции органического вещества, а также кальцификации служат энергетической и химической основой многих других сопряжелиых с ними процессов биогеохимического круговорота элементов, входящих в состав живого вещества и скелетной массы организмов, а также ряда элементов, динамика которых в водной среде связана с существованием в них редокс-зон. В числе элементов, которые входят в состав живого вещества, биогенные элементы; азот, фосфор, железо, марганец и кобальт. Динамика азота и фосфора в экосистеме рифа была рассмотрена нами выше. Здесь можно привести лишь ориентировочные расчеты масштабов их круговорота, основанные на оценках интенсивности биосинтеза живой биомассы в сообществах. Согласно Редфилду, молярное отношение С:N:Р в живом веществе близко к 100:16:1. Фактически же ввиду доминирования полузамкнутого никла биогенов в растительных ассоциациях рифов указанное соотношение выражается в них примерно так: 100:3:0,15 (Atkinson, 1981). В связи с этим при средней первичной продукции фотосинтеза, равной 4 г С/м² в сутки, интенсивность потока азота и фосфора должна выражаться соответственно величинами 120 и 15 мг/м² в сутки. При соотношении углерода, железа и марганца в биомассе водных растений 100:0,3:0,03 (по весу) поток железа, сопряженный с автотрофной продукцией, будет близок к 12 мг/м² и марганца — около 1 мг/м² в сутки. Поток кобальта может быть связан в сообществах рифа лишь с биосинтезом бактериальной биомассы (Сорокин, 1972). При средней суточной продукции бактериальной биомассы в экосистеме рифа около 0,5 мг С/м² суточный поток кобальта будет близок к 100 мкг/м².
Определений интерес представляет оценка глобальных масштабов биогеохимического метаболизма углерода и сопряженных с ним перечисленных выше элементов в сообществах коралловых рифов. Суммарная их площадь в Мировом океане оценивается в 6·10⁵ км², что составляет менее 0,2% площади всего Мирового океана и около 15% площади его мелководной зоны с глубинами 0-30 м (Smith, 1978). Валовая годовая первичная продукция (за вычетом дыхания самих растений-первопродуцентов) в сообществах рифов составляет в среднем около 1,5 кг С/м² - В этом случае глобальная первичная продукция коралловых рифов выразится величиной около 9·10⁸ тонн С,что составит 1,5-2% от суммарной первичной продукции фитопланктона в океане. При оценке этой величины следует учесть, что она в несколько раз превосходит суммарную годовую первичную продукцию всех высокопродуктивных зон апвеллинга, вместе взятых (Muscatine, 1980).
Исходя из приведенной выше ориентировочной величины глобальной продукции фотосинтеза в сообществах рифов, можно вычислить годовые величины глобальных потоков сопряженных с углеродом биогенных элементов. Поток азота в этом случае выразится величиной 2,7·10⁷ т, фосфора — 1,3·10⁶ т, железа — 3·10⁶ т и марганца — З·10⁵ т.
Сообщества коралловых рифов являются одним из главных генераторов карбонатных осадков на планете. Интенсивность этого глобального процесса можно оценить величиной около 2·10⁹ т СаСО₃ в год. Вполне естественно, что сообщества коралловых рифов, которые выполняют столь масштабную садку углекислого кальция из морской вода, служат также и важнейшим буфером, регулирующим концентрацию углекислоты в атмосфере. Кальцификация, протекающая на рифах, приводит к выведению части углекислоты из гидросферы и атмосферы. Тем самым биогеохимическая деятельность сообществ коралловых рифов снижает вероятность развития "парникового эффекта", который, как полагают, может быть вызван ростом концентрации CO₂ в атмосфере и может иметь катастрофические последствия прежде всего для будущего существования самих коралловых рифов. Повышение температуры у поверхности планеты как следствие "парникового эффекта" может вызвать таяние полярных льдов и столь быстрое повышение уровня океана, что коралловые рифы могут "утонуть", будучи не в состоянии догнать в процессе роста его поверхность.