На многих атоллах, располагающихся в продуктивных водах в зонах апвеллингов и дивергенций, существенным источником поступления биогенов и особенно фосфора является гуано рыбоядных птиц, которые образуют на таких островах значительные колонии. Концентрация фосфатов в воде может превышать на таких рифах 1 мкг ат./л (Христофорова, 1985). На некторых из островов древние рифы содержат залежи фосфоритов, возраст которых составляет 200-800 тыс. лет (Veeh, 1985). Такие фосфоритные руды в большом количестве имеются на островах Науру и Рождества. В малых количествах фосфориты встречаются в теле рифов многих атоллов. Вымываясь из этих рифов в процессах их эрозии, фосфаты с грунтовыми водами поступают в воды рифа, создавая в них повышенную концентрацию минерального фосфата (Gordon et al., 1971). Важную роль в поставке биогенов из их депо в донных осадках в толщу вод коралловых рифов играет постоянное их взмучивание на мелководьях рифа под влиянием волнения (Fanning et al., 1982; Entsch et al., 1983). В некоторых регионах одним из существенных путей поставки биогенов может являться ветровой нагон плавающих в поверхностном слое масс синезеленых водорослей Trichodesmium с окружающей рифы акватории открытого моря (Сорокин и др,, 1982). Перечисленные выше процессы создают постоянный поток солевых и органических форм биогенов в экосистему рифа. Потребление биогенов из этого потока, их удержание и регенерация из органических форм обеспечиваются включением биологических механизмов, участвующих в поставке и круговороте азота и фосфора.
Большое значение в процессах поставки и круговорота биогенов в сообществах рифа имеют перифитон и микрофитобентос. Сообщества перифитона, необычайно пышно развитые во всех донных биотопах рифа, содержат в своем составе водоросли типа мелких макрофитов, нитчаток, кораллин, колоний синезеленьх., богатую перифитонную микрофлору и простейших. Сообщества микрофитобентоса, обрастающие песчинки реальных осадков, также содержат в своем составе водоросли (диатомеи и синезеленые) и гетеротрофную микрофлору. Этим сообществам, содержащим в одной слизистой структуре и автотрофов и гетеротрофов, свойствен полузамкнутый тип биогенного обмена, при котором лишь небольшая доля необходимых биогенов поставляется извне, а остальные потребности организмов сообщества в биогенах удовлетворяются путем регенерации их солевых. форм из органических внутри самого сообщества. Значительная часть биогенов, которые образуются при деструкционной их регенерации гетеротрофными организмами, тут же, не покидая сообщества перифитона, усваивается автотрофами (Johannes et al., 1972). Такая внутренняя регенерация снижает потребность сообществ перифитона и микрофитобентоса в поставке в них биогенов из внешней среды. Особенно это касается азота, поскольку синезеленые водоросли и азотфиксирующие бактерии, обильно заселяющие эти сообщества, снабжают их связанным азотом. Поэтому в опытных условиях в этих сообществах почти постоянно наблюдается отдача биогенов в воду (Hanson, Gundersn, 1976). Что же касается фосфатов, то нашими опытами с меченым фосфором (см. табл. 5.5) было показано, что сообщества перифитона потребляют минеральный фосфат и одновременно отдают часть его в окружающую среду. В некоторых образцах перифитона такой обмен биогенами со средой у перифитонных. обрастаний рифа вообще практически незаметен, что указывает на высокую степень замкнутости в них биогенного цикла (Pomeroy et al., 1974).
Основную роль перифитонньх обрастаний и микрофитобентоса в биогенном цикле экосистемы рифа можно видеть в поставке в нее азота за счет азотфиксации, а также в усвоении микрофлорой сообществ перифитона органических биогенов из вод океана, проходящих над рифом. Наши исследования показали, что перифитонная микрофлора рифа использует для своего роста преимущественно органическое вещество вод океана, в состав которого входят азот и фосфор. Таким путем перифитонные сообщества способствуют поставке внешних биогенов из вод океана в экосистему рифа. Что же касается биогенного питания микрофитобентоса донныхосадков рифа, то значительную долю необходимых биогенов он получает за счет гетеротрофной их регенерации в толще осадка и за счет азотфиксации входящих в его состав в большом количестве синезеленых водорослей (Enhsch et al., 1983).
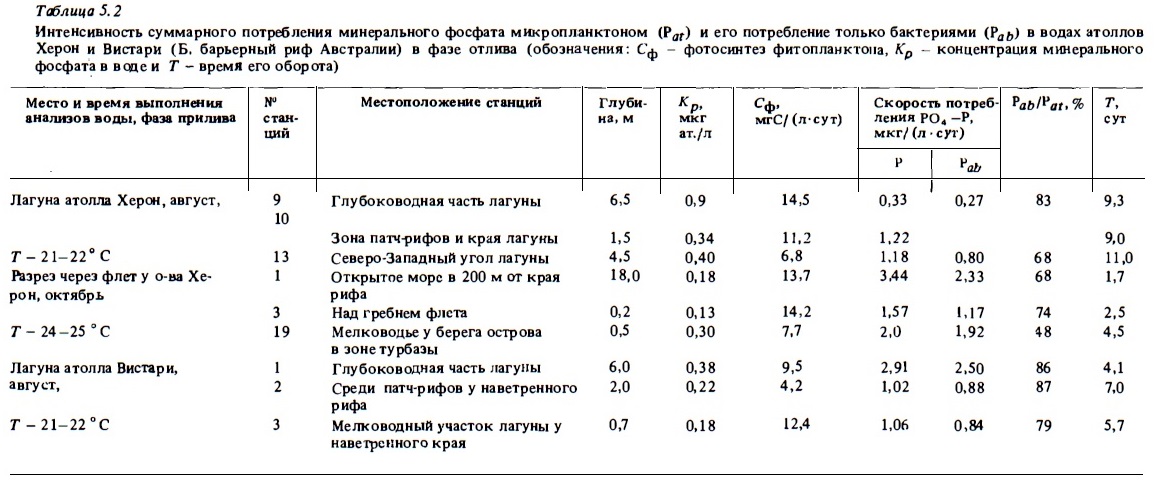
Солевые биогены потребляются из водной толщи также и макрофитами. Потребление происходит через поверхность их талломов (Pilson, Betzer, 1973; Nelson, 1985). Макрофиты способны запасать биогены впрок (Robishaux, 1985). Потребление биогенов можно заметить по убыли фосфата при прохождении воды через заросший ими флет (Webb et al., 1975). Однако даже при таких полях макрофитов на флете наряду с ними обитает достаточно много гетеротрофов, которые, выделяя фосфат, маскируют его потребление растениями. По расчету поток фосфатов из водной толщи в донные биотопы рифа с преобладанием макрофитов должен быть в дневное время близким к 0,1 мг ат./м2 в час. Прямые же измерения потока фосфора по изменению его концентрации в воде дают величину на порядок ниже. 0,012 мг ат./м2 в час (Pilson, Betzer, 1973). Морские травы потребляют биогены главным образом непосредственно из донных осадков с помощью своей корневой системы (Patriquin, 1972). Поэтому из заросли не могут оказывать заметного влияния на концентрацию солевых биогенов в проходящих над ними водах.
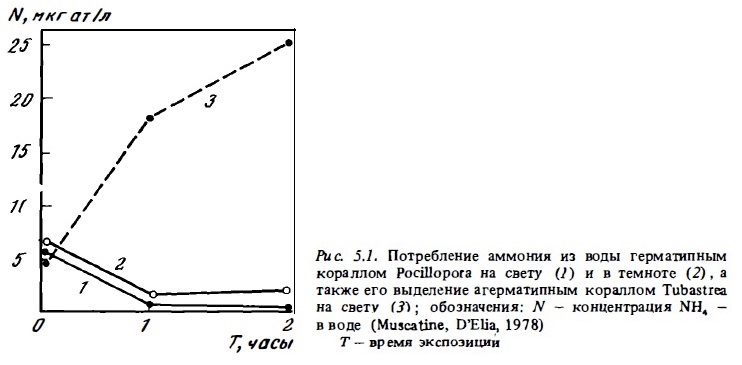
Многочисленными исследованиями доказана способность симбиотических животных рифа кораллов, горгонарий и тридакн потреблять солевые биогены при их концентрациях, близких к естественным: 0,3-1 мгк Р/л, 0,4-2 мкг ат./л О3 и 0,8-5 мкг/л РО4. Поглощение биогенов кораллами было установлено как по убыли их концентрации (Kawaguti, 1953; Webb, Wiebe, 1978; Muscatine, D’Elia, 1978; Пропп, 1981) (см. рис. 5.1), так и по потреблению ими солевых форм азота и фосфора, меченных изотопами: Р3 2, Р3 3 и N1 5 (Pomeroy et al., 1974; Sorokin, 1973; D'Elia, 1977; Burris, 1983; Wafar et al., 1985). Аналогичным образом ведут себя и тридакны, также содержащие в ткани своей мантии зооксантеллы (Burris, 1983; Propp, 1981), Способность к потреблению герматипными кораллами и тридакнами солевых форм биогенов непосредственно обусловлена наличием у них симбиотических водорослей зооксантелл. У кораллов с пониженным содержанием зооксантелл способность потреблять солевой азот резко снижается, а при их полном отсутствии прекращается вовсе. Агерматипные кораллы, не содержащие зооксантелл, такие, как Tubastrea, не поглощают солевых форм азота, а только выделяют их, как все другие животные, не имеющие растительный симбионтов (Muscatine et al., 1979; Muscatine, 1980). Зависимость интенсивности потреблении кораллами ионов аммония, нитратов и фосфатов от их концентрации в среде выражается кривыми, близкими к кинетической кривой Михаэлиса-Ментена (D’Elia, 1978; Webb, Wiebe, 1978; Muscatine, D'Elia, 1978) . Сам по себе характер этой зависимости, по мнению некоторый авторов, говорит в пользу того, что ионы биогенов потребляются кораллами из внешней среды с использованием механизма активного транспорта путем их переноса через клеточную мембрану с помощью специальных ферменгов — переносчиков пермеаз. Некоторое отличие кривых потребления кораллами ионов солевых биогенов от классической кривой Михаэлиса-Ментена указывает на то, что при их потреблении возможна и прямая диффузия внутрь клеток (Muscative, 1980).
Поступившие в клетки эпителия полипов кораллов ионы нитратов захватываются затем зооксантеллами и восстанавливаются в них до аммония (Crossland, Barnes, 1977а), включаясь в состав глютамина с помощью фермента глютамин-синтетазы. Затем через механизмы переаминирования образовавшаяся аминогруппа используется клетками зооксантелл для синтеза собственный аминокислот. Значительная доля произведенных таким путем зооксантеллами аминокислот транслоцируется в клетки полипа-хозяина (Muscatine, Cernichari, 1969; Trench, 1971; Lewis, Smith, 1971; Muscatine, 1980). Аккумуляция органических форм азота, помеченного изотопом N1 5, в тканях кораллов, которые инкубировались в среде, обогащенной меченным N1 5 нитратом, была доказана прямыми опытами (Marian, 1979).
Обратная поставка конечных продуктов минерализации аминокислот из клеток полипа-хозяина в клетки симбиотических зооксантелл была доказана в опытах с актиниями Anthopleura (Marian, 1979), которым скармливался белок, содержащий N1 5 в составе азота аминогрупп. В изолированных из их тканей зооксантеллах в конце экспозиции обнаруживалась метка N1 5. Таким путем у кораллов и у других обитателей рифа, имеющих растительные симбионты, образуется замкнутый цикл основных биогенных элементов азота и фосфора, благодаря которому существенно снижаются их потери в окружающую среду.
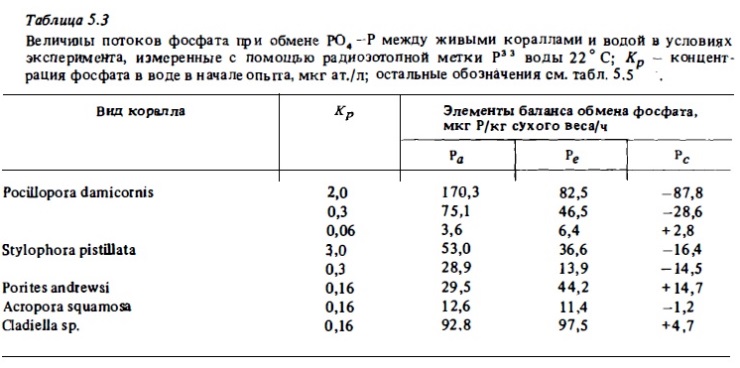
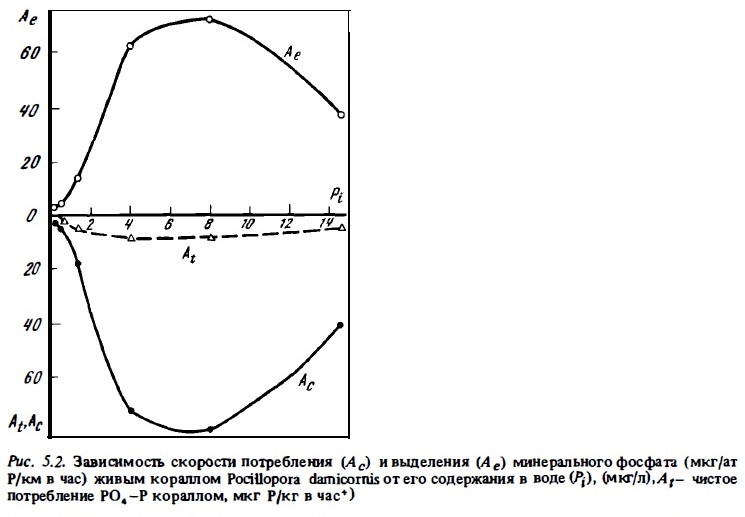
Значимость представленных выше результатов исследований потребления кораллами минеральных форм биогенов, основанных на прямом определении убыли их концентрации в присутствии коралла, для количественной оценки баланса биогенов весьма условна. Дело в том, что, несмотря на определенную замкнутость обмена биогенов и симбиотических животных, они одновременно с потреблением их солевых форм выделяют их во внешнюю среду в процессе обмена при участии других ферментных систем. Поэтому при изменении убыли концентрации солевого биогена в среде в присутствии коралла фактически измеряют не абсолютное его потребление, а некоторую меньшую неопределенную величину, отражающую равновесие процессов потребления и выделения и близкую к разности этих разнонаправленных потоков биогенов. Оценка истинной скорости потребления минеральных форм биогенов кораллами возможна лишь с применением меченных изотопами их форм (Pomeroy et al., 1974; Burris, 1983). Мы пытались измерить истинные скорости потоков фосфатов у кораллов при потреблении и выделении ими минерального фосфата, использовав для этой цели радиоизотоп Р3 3.
Результаты опытов показали, что при концентрациях РО4—Р в воде, близких к естественным (0,1—0,3 мкг ат./л), изменения абсолютной концентрации РО4—Р в воде вообще не отражают истинной скорости его потребления кораллами, поскольку потребление РО4—Р в этих условиях в значительной мере нивелируется его одновременным выделением. Часто выделение РО4—Р перекрывает его потребление. При этом наблюдается не убыль, а, наоборот, прирост фосфата в воде в присутствии коралла (табл. 5.3, рис. 5.2). При повышенных концентрациях фосфата в опытных аквариумах результаты определений потребления фосфата кораллами из воды по изменению его абсолютной концентрации занижаются в 2-10 раз. Общая интенсивность потребления фосфата кораллами в темноте обычно была ниже, чем на свету. Интенсивность выделения ими РО4—Р в темноте несколько возрастает (рис. 5 .4) и, таким образом, уравновешивает его потребление. В этих условиях изменения концентрации фосфата в воде за время опыта практически не происходит, хотя интенсивность потоков в темноте фактически остается почти такой же, как на свету (см. рис.5.2, 5.4).
Наряду с выделением минеральный биогенов кораллы выделяют в среду органические соединения азота и фосфора (Johannes, Webb, 1970; Muscatine, D’Elia, 1978). Чистая отдача азота и фосфора кораллами и другими симбиотическими животными рифа намного ниже, чем у обычный животных, не имеющих в своих тканях симбиотических водорослей (Lewis, D. 1973; Muscatine, Porter, 1977). Такой полузамкнутый обмен биогенов у кораллов способствует удержанию основного их запаса в пределах экосистемы рифа. Основным источником его пополнения является гетеротрофное питание кораллов. Потребление ими солевых биогенов имеет подчиненное значение, компенсируя отдачу биогенов в воду (Пропп, 1981).
В числе биологических механизмов поставки в экосистему рифа азота и фосфора из проходящих над ним вод океана следует также указать на потребление органической взвеси (в основном бактериопланктона и фитотанктона) донными и планктонными фильтраторами рифа из проходящих над рифом вод океана). Органические формы биогенов, содержащиеся в водах океана (см. табл. 5.1), потребляются планктонными бактериями, затем поставляются в экосистему рифа через их выедание животными-филттраторами и седиментаторами. Этот путь поставки был продемонстрирован нами (Sorokin, 1973) прямыми опытами, из которых видно, что кораллы усваивают меченый минеральный фосфат через бактериальное звено интенсивнее, чем непосредственно в виде РО4-Р. Важное значение в поставке биогенов играет также выедание зоопианктона, поступающего с проходящими над рифами водами океана, хищниками рифа, в основном рифовым демерсальным хищным мезо- и макрозоогтанктоном, кораллами и рыбами. Мелкие рыбы-планктофаги, а также танктонные личинки многих донных рыб рифа постоянно выедают зоошанктон из проходящих над рифом вод, поставляя азот и фосфор в донные биотош рифа в виде фекального материала, который подвергается в них минерализации через пищевую цепь поедающими его донные беспозвоночными, включая сами кораллы (Meyer et аl., 1983).