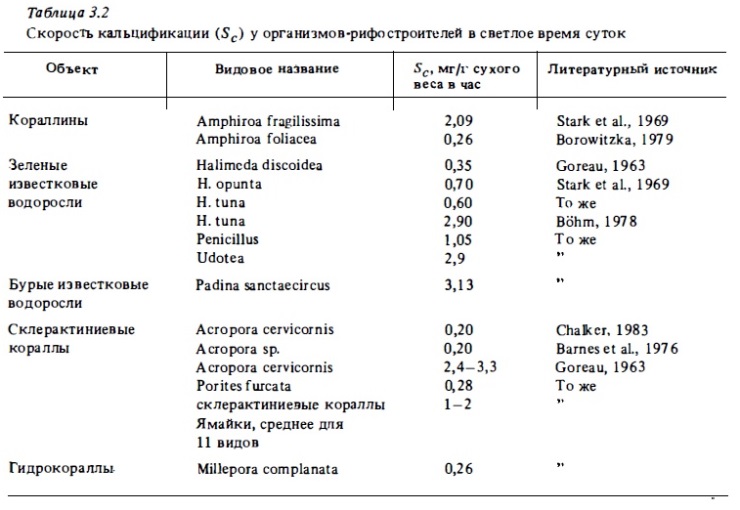
Скорость кальцификации у кораллов по результатам измерений с помощью Са4 5 варьирует в пределах от 0,2 до 3,3 мг/г в час. Данная величина (3,3 г) была получена автором радиоизотопного метода Горо. Впоследствии его данные подверглись критике (Borowitzka, 1979). Оказалось, что в начале экспозиции колонии коралла в присутствии меченого кальция его потребление проходит в значительной мере за счет обмена ионами Са2+ между средой и кристаллами арагонита в скелете животного, а не за счет продукции нового карбоната кальция. В результате определяемая скорость кальцификации оказывается завышенной, что, по-видимому, и характерно для данных, проводимых в работах Т. Горо. Последние определения показали, что скорость кальцификации у большинства изученных видов кораллов варьирует в пределах 0,2-0,6 мг/ (г·ч), что должно соответствовать годовой продукции 2—10 кг/м2 СаСО3 при среднем покрытии площади рифа кораллами 10—30%.
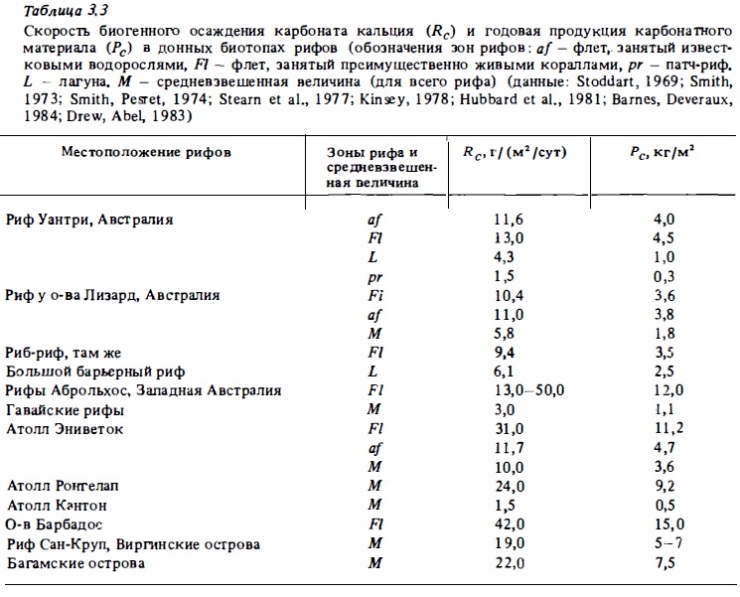
Многочисленные измерения суммарной продукции карбонатного материала в донных биотопах коралловых рифов показывают, что наибольших величин она достигает на риф-флете независимо от того, заселен он кораллами или известковыми водорослями ("algal ridje", "algal pavement") . Как видно из табл.3.3, в которой суммированы результаты этих измерений, на риф-флете за сутки кораллами и кораллинами отлагается более 10 г карбонатного материала на 1 м2. Эта величина оказывается соизмеримой, а иногда и превышает продукцию органического вещества в донных биотопах коралловых рифов, которая составляет в зоне флета 8—12 г за сутки в расчете на сухой вес. Весьма значительна скорость кальцификации в лагуне, где она осуществляется известниковыми водорослями и фораминиферами (4 г/м2). Средние величины биогенной продукции СаСО3, определяющие всего сообщества ряда рифов и атоллов, дали величины от 4 до 10 г/ (м2·сут), или от 1 до 15 кг/ (м2·год) (Smith, Kinsey, 1976; Glynn, 1977; Stearn et al., 1977). Рекордно высокая скорость кальцификации — около 50 г СаСОз — была зарегистрирована на флете высокогорных рифов Аброльхос в Западной Австралии (Johannes et al., 1983). Средняя годовая продукция СаСО3 на этих рифах составляла около 12 кг/м2, что соответствует скорости роста рифа (учитывая пористость) около 2 см в год. На флете и в биотопах внешнего склона рифа годовая продукция карбонатного материала оценивается в 3-4,5 кг/ (м2·год) (Smith, 1983). Эти данные показывают, что экосистемы рифов являются мощными генераторами карбонатного материала. За год на 1 км2 они продуцируют около 2-4 тыс. тонн биогенного карбоната, в состав которого наряду с кальцием входит в значительном количестве магний (5-10%). Рифовые карбонаты содержат также стронций, железо и ряд тяжелых металлов (Fe, Си, Ni, Cd, Zn, РЬ, John, 1974). Суммарная площадь коралловых рифов составляет 6 · 105 км2. Соответственно годовую продукцию производимого ими карбонатного материала можно оценить примерно в 2,5 млрд т.
Скорость линейного роста кораллов в значительной мере определяется скоростью кальцификации. Поэтому факторы, влияющие на кальцификацию, влияют на скорость их роста, а также и на форму их колоний, поскольку при изменении скорости кальцификации, сохранении роста мягких тканей и числа полипов меняются соотношения между гаощадью поверхности коралла и числом полипов (Goreau, 1963). В числе факторов, влияющих на скорость кальцификации, следует упомянуть обсуждавшиеся выше условия освещенности, а также антропогенную эвтрофикацию и влияние штормов. В условиях эвтрофирования повышенная концентрация биогенов в воде может тормозить кристаллизацию СаСО3 (Simkiss, 1964). В условиях эвтрофикации возрастает мутность воды, что ухудшает освещенность и тормозит светозависимую кальцификацию кораллов (Dodge, Vaisnys, 1977; Dodge, 1981). Воздействие ураганов на риф приводит прежде всего к ломке крупных зрелых колоний кораллов, которые имеют большую площадь соприкосновения с волновым стрессом и в большей степени ослаблены сверлящими организмами. На месте этих сломанных медленно растущих колоний поселяются интенсивно растущие молодые кораллы. Таким образом, происходит постоянное омоложение рифостроящих сообществ кораллов под действием штормов и ураганов, в результате чего общая интенсивность кальцификации на таких рифах возрастает (Highsmith et al., 1980). Замечено, что на барьерном рифе скорость линейного роста кораллов по мере удаления от берега снижается (Ladale, 1983).
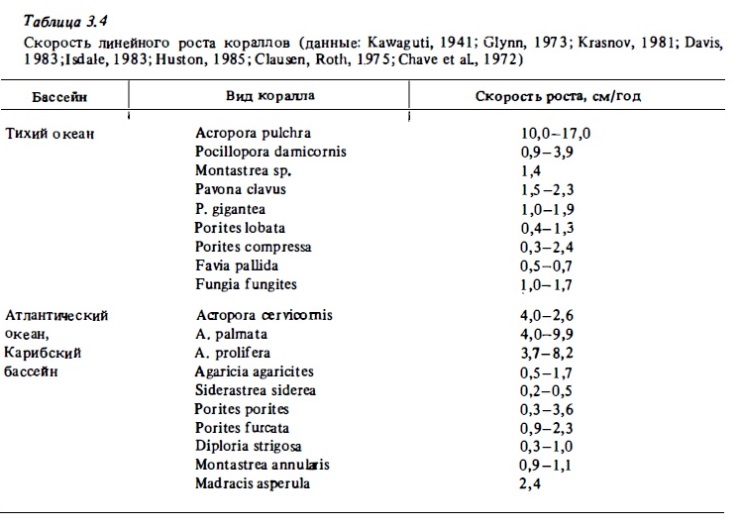
Показателем интенсивности кальцификации кораллов может быть и скорость их линейного и весового роста. Результаты их измерений разными авторами представлены в табл. 3.4. Согласно этим данным, наибольшая скорость роста характерна для оппортунистических и наиболее массовый видов кораллов с ветвистой формой колоний. В числе наиболее быстро растущих кораллов массовые виды ветвистых кораллов: Acro-pora cervicornis, А. palmata, А. forrnosa. Скорость их линейного роста составляет от 4 до 26 см в год (Huston, 1985). Скорость роста ветвистых кораллов Pocillopora, Porites близка к 1—4 см в год. У массиннгх форм Porites, Montastrea она составляет 0,3-2,5 см в год (Krasnov, 1981; Поляков, 1982; Crossland, 1981; Alcala et al., 1981). Скорость линейного роста различна в разных частях одной и той же колонии и у разных колоний одного вида (Stephenson, Stephenson, 1933; Goreau, Goreau, 1959; Buddemeyer et al., 1974). Эта разница у некоторых кораллов, например у Manicina areolata, может превышать порядок величин (Goreau, 1961). Существенно, что ионы кальция могут транспортироваться по ценосарку колонии коралла в зоны наиболее интенсивной кальцификации (Taylor, 1977).
Как видно из табл. 3.4, представители одного и того же рода кораллов в разных регионах растут с разной скоростью. Более того, линейная скорость роста кораллов заметно варьирует и в разные сезоны года. Эти колебания особенно велики у кораллов на рифах., расположенных у границ тропической зоны, где значительны сезонные изменения температуры (Sblnn, 1966; Glynn, 1977). На рифах Абральхос, расположенных за пределами тропической зоны на 28° ю. ш., коралл Acropora formosa летом рос в 5 раз быстрее, чем зимой (Crossland, 1981). В опытах с переносом кораллов в меняющиеся температурные условия, а также путем наблюдений за сезонным ростом кораллов был найден оптимум температуры для их роста, который близок к 26-27°С (Уар, Gomez, 1981). При температуре ниже 16° он прекращался (Wells, 1957; Buddemeyer, Kinzie, 1976; Weber, White, 1977). Температура выше 32° является для гавайских кораллов летальной (Jokiel, Coles, 1977). Температурный оптимум для процесса кальцификации кораллов в лабораторных условиях был близок к 27-30°С (Glynn, Stewart, 1973; Clau-sen, Roth, 1975).
Как уже отмечалось выше, важным фактором, влияющим на рост кораллов, являются условия освещенности. Исследования показали, что уже в пределах глубин от 5 до 20 м скорость линейного роста кораллов заметно снижается вместе с падением освещенности, хотя и у самой поверхности может наблюдаться некоторое его уменьшение, связанное с ингибирующим эффектом избыточного света (Buddemeyer et al., 1974; Bak, 1976; Smith, Harrison, 1977; Highsmith 1979; Roth et al., 1982). Снижение скорости роста кораллов с глубиной происходит, несмотря на наличие у кораллов определенных физиологических механизмов световой адаптации. Например, коралл Porites lutea на глубине 4 м рост в 2,5 раза быстрее, чем на глубине 30 м (Buddemeyer et al., 1974). Связано это, по-видимому, с тем, что свет, как и температура водаг, не является единственным меняющимся с глубиной фактором, который может контролировать рост кораллов. В числе таких факторов можно назвать силу движения воды, волнобой, концентрацию взвеси планктона как источников гетеротрофного питания кораллов.
Линейная вертикальная скорость роста самих рифов определяется геологами достаточно точно на основе радиоуглеродный датировок кернов. До достижения современными рифами поверхности моря в период голоценовой трансгрессии скорость роста их скального фундамента составляла 1-12 мм в год. При таком годовом приросте продукция карбонатного материала сообществом герматипов рифа даже без учета одновременно идущей эрозии выражалась величинами 3-20 г/ (м2·год), что в целом несколько выше оценок величины его продукции на современных рифах, достигших поверхности моря. Скорость роста рифов, равно как и состав организмов-рифостроителей, в большей степени зависит от воздействия волнения. В условиях сильного волнобоя, такого, как, например* на атоллах пассатной зоны Тихого океана, рост островов идет в значительной мере за счет известковых водорослей кораллин и в меньшей степени за счет кораллов. В этих условиях рифы растут медленнее, в большей степени консолидированы, чем рифы Атлантики с преобладанием на них кораллов как основных рифостроителей (Adey, 1978).
Консолидированный биогенный карбонат, который производится на флете современных рифов, достигших поверхности моря, частично подвергается эрозии и перерабатывается волнобоем в рыхлый материал, который сносится в лагуну или на внешнюю сторону рифа. Стекая по склонам скального основания рифа, этот материал участвует в формировании склоновых карбонатных осадков дна окружающего рифы моря. Определение скорости седиментации рыхлого карбонатного материала в лагунах производилось разными методами: по балансу карбонатов (риф Уантри, Davies, 1977), по радиоуглеродным датировкам проб, взятых под дном лагун (рифы Большого барьера в Австралии, Frankel, 1977), по выделению карбоната с фекалиями животных, питающихся кораллами и донными осадками (Bardach, 1961). Эти определения дали сходные результаты: 1-3 мм в год.