
Основными источниками кислорода для воды являются фотосинтезирующие растения и атмосфера. В атмосфере содержится около 21% кислорода. В холодной воде, находящейся в контакте с воздухом атмосферы, в состоянии насыщения растворено не более 15 мг/л кислорода, т. е. массовая доля кислорода составляет примерно 0,0015. Столь малые концентрации затрудняют извлечение кислорода водными организмами и требуют строгого контроля за содержанием кислорода в воде.
Содержание кислорода в воде, насыщенной воздухом, зависит от температуры и солености (хлорности). При постоянной температуре концентрация насыщения воды кислородом линейно связана с хлорностью. Однако с увеличением температуры наклон кривой уменьшается (рис. 4.1). Зависимость концентрации насыщения от температуры при постоянной хлорности, если ее изобразить на полулогарифмической координатной бумаге, выглядит почти линейно.
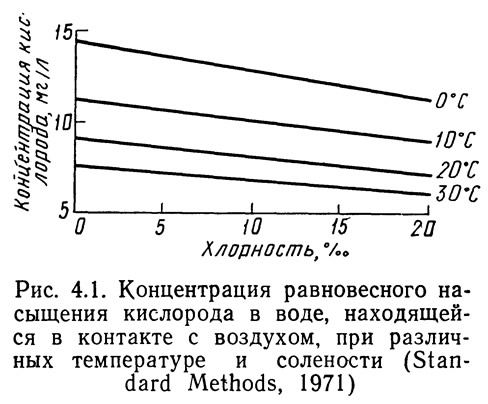
В табл. 4.1 показано, как концентрация насыщения воды кислородом зависит от температуры и солености. Из таблицы видно, что в холодной воде может содержаться больше кислорода, чем в теплой, а в пресной больше, чем в соленой. Иными словами, увеличение как солености, так и температуры приводит к уменьшению содержания кислорода в воде при 100 %-ном насыщении.
Скорость переноса кислорода из воздушной среды в водную зависит от температуры, солености и степени насыщения воды, а также от турбулентности поверхности раздела вода — воздух. Температура воды, соленость, степень насыщения — все эти факторы влияют на градиент концентрации, являющийся движущей силой процесса переноса кислорода. Турбулентность раздела вода — воздух значительно увеличивает площадь контакта двух сред: чем больше площадь поверхности раздела, тем больше скорость переноса кислорода. Перенос кислорода в воде почти целиком обусловлен циркуляцией, течениями и турбулентностью. Диффузия кислорода столь мала, что не имеет практического значения. Например, для увеличения концентрации кислорода на глубине 10 м от 0 до 0,4 мг/л диффузией потребовалось бы около 600 лет (Reid, 1961).
Градиент концентрации оказывает большое влияние на скорость переноса кислорода через поверхность раздела вода—воздух. Поскольку содержание кислорода в атмосфере может лишь незначительно отклоняться от 21%, скорость переноса определяется отношением данной концентрации кислорода в воде к концентрации насыщения.. Математически градиент концентрации можно записать в виде
где Cs — концентрация кислорода при 100 %-ном насыщении; Сt — концентрация кислорода в момент времени t; ΔСt — градиент концентрации в момент времени t.
Скорость переноса кислорода с увеличением градиента концентрации возрастает, поэтому по мере приближения содержания кислорода в воде к 100 %-ному насыщению скорость переноса уменьшается.
Именно поэтому значительно труднее полностью насытить воду кислородом, чем повысить концентрацию на 1—2 мг/л.
Математически перенос кислорода можно описать следующим уравнением:
где ΔС/Δt — изменение концентрации за время Δt; К — константа, учитывающая влияние турбулентности и зависящая от условий; А — площадь поверхности контакта вода—воздух; Cs и Ct — те же, что и в уравнении (4.1).
Уравнение (4.2) чаще встречается в дифференциальной форме
Это уравнение показывает, что скорость изменения концентрации dC/dt равна произведению константны К, учитывающей влияние турбулентности поверхности раздела вода—воздух, на площадь поверхности контакта и градиент концентрации Cs—Ct.
Фотосинтез — второй основной источник кислорода для водных систем. Кислород является побочным продуктом фотосинтеза и выделяется в воду фотосинтезирующими растениями. Однако фотосинтез и продуцирование кислорода могут происходить только при наличии света. Те водоросли, которые в дневное время выделяют кислород, ночью потребляют его. В солнечные дни количество кислорода, выделяемого фотосинтезирующими растениями, может быть достаточно для пересыщения поверхностных вод кислородом. В этом случае он выделяется в атмосферу. Фотосинтетическая продукция кислорода несколько отстает от суточного хода получаемой Землей солнечной энергии, поэтому содержание кислорода в естественных водоемах и прудах минимально на рассвете. Максимальные концентрации обычно наблюдаются в середине дня или несколько позднее. Таким образом, определение концентрации кислорода для выявления возможного его дефицита следует проводить на рассвете.
Пресноводные системы
Озера и пруды. Для водоемов, глубина которых превышает толщину слоя фотосинтеза, характерны некоторые особенности распределения кислорода. В верхнем слое воды продуцирование кислорода в результате фотосинтетической активности, осредненное за сутки, превышает его потребление. В нижележащих слоях кислорода производится меньше, чем потребляется. На некоторой глубине количество продуцируемого кислорода равно количеству потребляемого. Эта глубина называется глубиной компенсации. Ниже этой точки потребности кислорода на дыхание превышают его продуцирование, так как фотосинтез ограничен количеством проникающей сюда световой энергии. Кроме того, сюда же опускается отмерший планктон из вышележащих слоев. По мере погружения частицы планктона облепляются бактериями, которые извлекают из них органические компоненты. Кислород требуется для дыхания бактерий и других обитателей этих глубин.
Вертикальное распределение кислорода в озерах и прудах зависит от циркуляции и потенциальной продукции вод. На рис. 4.2 представлено типичное вертикальное распределение кислорода в эвтрофном высокопродуктивном озере. Зимой при наличии ледового покрова содержание кислорода незначительно убывает с глубиной и со временем медленно уменьшается во всей толще вод. Непосредственно у поверхности концентрация может быть несколько выше, что обусловлено фотосинтетической активностью. Однако фотосинтез лимитируется количеством света, проникающего сквозь толщу льда, а также низкой температурой воды. С наступлением весны и таянием льда начинается обмен кислородом между водной средой и атмосферой. Одновременно начинается весеннее перемешивание вод. Исчезновение ледового покрова и перемешивание приводят к тому, что содержание кислорода становится довольно высоким и одинаковым по всей глубине. С приближением лета происходит стратификация водной толщи. Воды, находящиеся над термоклином, перемешанные и прогретые, благодаря активному фотосинтезу и обмену с атмосферой характеризуются высоким содержанием кислорода. Поверхностные воды могут даже в течение дня достигать насыщения. Воды термоклина и нижележащие не контактируют с атмосферой из-за температурной стратификации. Сюда в значительном количестве поступают органические остатки погибших в вышележащих продуктивных слоях организмов. Температура этих вод несколько выше, чем зимой. Это способствует интенсификации метаболизма бактерий. Процессы разложения и дыхания приводят к быстрому использованию получаемого кислорода.
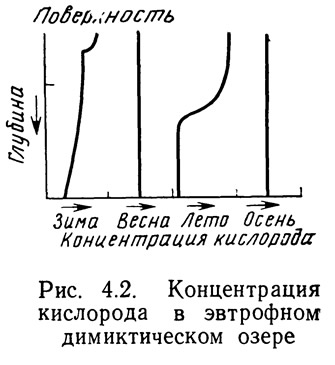
Часто к концу лета воды ниже термоклина становятся анаэробными. В анаэробных условиях возможно накопление (метана и сероводорода. Эти газы с характерным неприятным запахом токсичны для обитателей водоемов. С наступлением осени поверхностные воды охлаждаются, термоклин разрушается — начинается осеннее перемешивание. Концентрация кислорода выравнивается по всей глубине, а излишки углекислоты, сероводорода и метана высвобождаются в атмосферу.
Область быстрого снижения концентрации кислорода, называемая клиноградом1, обычно располагается вблизи от термоклина, так как вышележащие воды перемешиваются и хорошо вентилируются. Однако термоклин и скачок кислорода не всегда полностью совпадают по глубине. В эвтрофных водах клиноград обычно расположен на глубине термоклина или несколько выше, так как высокая продуктивность (большое число клеток фитопланктона) вызывает быстрое ослабление света. В олиготрофных, низкопродуктивных водах клиноград менее выражен и может вообще отсутствовать.
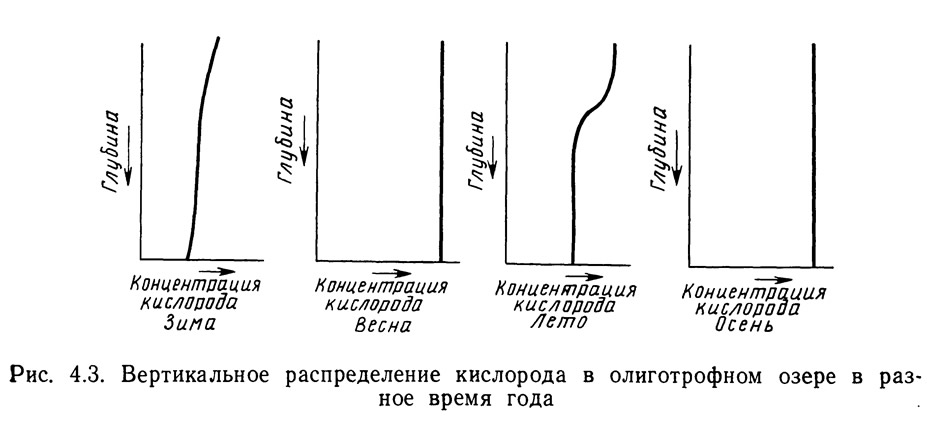
Олиготрофные низкопродуктивные воды относительно чисты и прозрачны. На рис. 4.3 показано распределение кислорода по сезонам в олиготрофном димиктическом озере. Кривые вертикального распределения очень сходны с кривыми для эвтрофного озера. Различия заключаются в том, что зимой, когда процессы дыхания и разложения органических веществ замедлены, концентрация кислорода в олиготрофном озере несколько выше, чем в эвтрофном. Весной и осенью распределение кислорода в олиготрофном озере такое же, как в эвтрофном. Наиболее значительные различия в распределении кислорода между этими двумя типами водоемов наблюдаются летом. В олиготрофном озере слой компенсации располагается на большей глубине в связи с меньшей продуктивностью и соответственно меньшим экранирующим эффектом планктона. Пониженная продуктивность требует также меньше кислорода на дыхание и разложение органических веществ. В результате летом наблюдается высокая концентрация кислорода выше клинограда (правда, несколько ниже, чем в эвтрофном озере), сравнительно слабо выраженный клиноград и медленное снижение в нижележащих слоях. В олиготрофных озерах анаэробные условия развиваются редко, даже к концу лета в глубинных водах.
Эвтрофные и олиготрофные воды были описаны в качестве экстремальных примеров. Кроме них существуют озера и пруды с промежуточными характеристиками. Рыбоводные пруды часто недостаточно глубоки для развития выраженного термоклина, но могут иметь довольно резко выраженный клиноград. Причиной его образования может быть избыточное кормление. При правильной биотехнике в прудах не должен образовываться заметный клиноград. При устройстве коллекторов, садков и других приспособлений для выращивания водных организмов необходимо исключать возможность обитания культивируемых объектов под слоем скачка кислорода. Это оборудование нельзя также размещать в местах, где возможно значительное выделение сероводорода под клиноградом, даже если оно установлено над ним, так как проходящие сквозь столб воды пузырьки сероводорода могут оказаться глубительными для культивируемых объектов. Кроме того, следует избегать размещения водозаборных устройств под клиноградом, если не обеспечена достаточная аэрация. Хотя аэрация обходится довольно дорого, в некоторых районах, возможно, целесообразно для получения охлажденной воды откачивать ее из-под клинограда.
Проточные воды. Проточные воды насыщаются кислородом в результате фотосинтетической активности и контакта с воздухом атмосферы. Однако фотосинтез не имеет здесь такого значения, как в озерах и прудах, поскольку в проточной воде рост фитопланктона затруднен. Чем больше скорость и турбулентность потока, тем меньшую роль играет фотосинтез как источник кислорода. Прикрепленные водоросли могут продуцировать значительное количество кислорода, особенно в реках и ручьях с медленным течением.
Содержание кислорода в проточных водных системах (реках, ручьях, рыбоводных каналах) зависит от скорости течения, расхода системы, мутности (в частности, обусловленной неживым веществом) и температуры воды, поперечного сечения ручья или канала и количества получаемой солнечной энергии. Влияние этих факторов проявляется косвенно: они определяют скорость фотосинтеза, дыхание, разложение органического вещества и интенсивность физической аэрации, т. е. содержание кислорода в системе. Относительное значение каждого из них определить довольно трудно, поскольку они в значительной степени различаются для разных систем и изменяются для каждой системы во времени и пространстве.
Годовой цикл кислорода в реках и ручьях тесно коррелирует с температурой. В большинстве рек содержание кислорода достигает максимума зимой, а минимума — летом. Однако эта основная зависимость может нарушаться несколькими факторами. Ледовый покров подавляет фотосинтез и уменьшает зимний максимум кислорода. Весеннее уменьшение концентрации кислорода может быть связано с выносом подводной растительности во время половодья. Минимум растворенного кислорода в конце лета часто обусловлен высокой температурой воды, уменьшением перемешивания и расхода или интенсивным разложением продуцированного за лето органического вещества. При достаточном содержании питательных веществ возможно быстрое размножение фитопланктона («цветение»). В это время содержание кислорода днем может резко увеличиваться.
Суточный цикл кислорода в ручьях в основном определяется флуктуациями температуры и соотношением процессов фотосинтеза и дыхания. В быстрых чистых ручьях) с ограниченным количеством растительности концентрация кислорода благодаря турбулентности поддерживается на уровне 100%-ного насыщения или около него. Фотосинтетическая продукция кислорода за светлое время суток часто превышает диффузию его в атмосферу и потребление на дыхание, вместе взятые. В результате днем происходит перенасыщение воды кислородом, а ночью концентрация его снижается до 100 %-ного насыщения. Зимой максимумы суточной продукции кислорода и температуры наблюдаются примерно в одно время. Летом увеличение продукции кислорода часто отстает от роста температуры на 2—3 ч. Это приводит к тому, что содержание кислорода остается высоким даже после наступления темноты. Минимум кислорода в ручьях этого типа наблюдается перед понижением температуры ранним утром, но точное его время зависит в определенной степени от потребления кислорода. Большое влияние на концентрацию кислорода в ручьях этого типа оказывает интенсивность солнечной радиации.
Распределение кислорода по течению реки зависит от многих факторов. В верхнем .течении концентрация кислорода в основном определяется температурой и турбулентностью потока. Ниже по течению, где река замедляет свой ход и появляется обильная растительность, содержание кислорода обусловлено главным образом процессами фотосинтеза и дыхания. Максимум кислорода наблюдается здесь в полдень, а минимум — ранним утром. В нижнем течении с возрастанием мутности и интенсификацией процессов разложения органического вещества концентрация кислорода уменьшается. Поступление загрязняющих веществ, а также подъем уровня вод в болотах и маршах приводят к уменьшению содержания кислорода в месте впадения рек и ниже по течению.
Содержание кислорода в той части реки, которая расположена ниже дамб и водохранилищ, зависит от того, с какого уровня водохранилища производится забор воды. Воды, забираемые из гиполимниона при развитой летней стратификации, бедны кислородом и могут оказать большое влияние на все компоненты экосистемы ниже по течению. Если вода забирается из верхнего слоя водохранилища, богатого кислородом, это может способствовать повышению содержания кислорода в нижнем течении реки.
Морские системы
Концентрация кислорода в естественных морских системах (океанах, морях) может изменяться от нуля, например в некоторых анаэробных областях, таких, как глубинные воды Черного моря, до равновесного насыщения и даже перенасыщения в водах, омывающих коралловые рифы2. Поскольку соленость океанских вод составляет приблизительно 35 %о, концентрация равновесного насыщения в океанах ниже, чем в пресноводных водоемах. Для поверхностных тропических вод эта концентрация составляет 6—7 мг/л, а для полярных может превышать 11 мг/л. Концентрация глубинных океанских вод, хотя они и не насыщены кислородом, часто превышает 7 мг/л (Raymont, 1963).
Хотя концентрация кислорода в различных океанах и даже в разных районах одного океана колеблется, можно обнаружить определенные закономерности в вертикальном распределении. На рис. 4.4 показана типичная кривая вертикального распределения кислорода для вод субтропической зоны океанов.
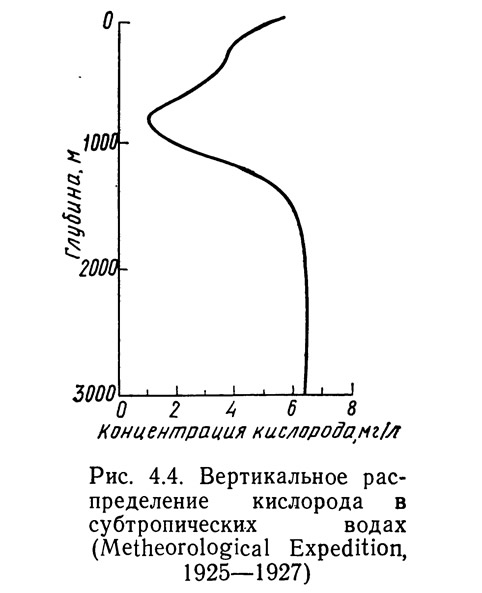
Концентрация кислорода в поверхностном слое в этих районах довольно высока. Поступление кислорода из атмосферы и в процессе фотосинтеза поддерживает концентрацию на уровне, близком к равновесному насыщению (5—7 мг/л). Ниже глубины слоя фотосинтеза концентрация резко убывает, образуя область, известную как «зона кислородного минимума». Глубина ее колеблется от 150 м в некоторых морях до 1000 м в океанах. Концентрация кислорода в этой зоне может быть ниже 1,4 мг/л. Наиболее низких значений (0,14 мг/л) она достигает в зоне кислородного минимума у берегов Южной и Центральной Америки в Тихом океане. Концентрация кислорода в зоне кислородного минимума в Атлантическом океане, по-видимому, в среднем несколько выше, чем в Тихом. Это различие может быть объяснено смешением больших количеств подповерхностных вод с Тихоокеанскими промежуточными водами, что приводит к образованию верхнего минимума кислорода с очень низкими концентрациями в юго-восточной части Тихого океана.3
Ниже зоны кислородного минимума с увеличением глубины концентрация кислорода возрастает и в глубинных водах достигает довольно высоких значений (Raymont, 1963).
Образование зоны кислородного минимума, по-видимому, обусловлено несколькими причинами. В областях минимальных концентраций скорости течений, вероятно, достигают минимальных значений. Поскольку термоклин располагается на этой же глубине, происходит быстрое увеличение плотности и вязкости воды. В результате скорость опускания детрита уменьшается, что приводит к увеличению концентрации разлагающегося органического вещества. Для его окисления требуется значительное количество кислорода. На глубинах, где прослеживается зона кислородного минимума, обнаруживаются большие скопления зоопланктона, потребляющего кислород в процессе метаболизма. Совместное действие всех перечисленных выше факторов приводит к уменьшению концентрации растворенного кислорода в воде на данных горизонтах.
Хотя описанное выше вертикальное распределение дает представление о характере изменения концентрации кислорода с глубиной, кривые вертикального распределения в некоторых районах океана отличаются от типичных. В полярных областях, где происходит погружение поверхностных вод, содержание кислорода остается почти постоянным во всей толще. В таких районах подъема вод, как, например, у западных берегов Северной и Южной Америки концентрация кислорода на поверхности или вблизи нее значительно ниже концентрации равновесного насыщения. Поверхностные слои в высокопродуктивных районах, например к востоку от Гренландии или около некоторых коралловых рифов, в периоды интенсивной фотосинтетической деятельности могут быть перенасыщены кислородом.
Солоноватоводные системы
Концентрация кислорода в эстуариях колеблется в широком диапазоне и зависит от многих факторов. Поскольку степень насыщения кислородом уменьшается с увеличением температуры и солености, градиент кислорода часто расположен вдоль эстуария с максимальными концентрациями в верхней по течению части эстуария. В периоды большого речного расхода и притока пресной воды концентрация кислорода в эстуарии увеличивается. В сухое время года с повышением солености эстуарных вод следует ожидать снижения концентрации растворенного кислорода.
Концентрация кислорода в водах эстуария может определяться стратификацией, глубиной, наличием растительности, приливами, временем года и даже дня. При плотностной стратификации в эстуарии может существовать значительная разница в концентрации кислорода между поверхностными и глубинными водами. Например, в Чесапикском заливе концентрация кислорода летом в поверхностных водах составляет 90—100% равновесного насыщения, в то время как в придонных — 40—50% (Raymont, 1963). Это свидетельствует о продуцировании кислорода в процессе фотосинтеза в верхних слоях и об утилизации его при разложении органических веществ или о присутствии бедных кислородом морских вод в нижних слоях.
Суточные и сезонные колебания концентрации кислорода в эстуариях зависят от происхождения пресноводного источника, морфологии бассейна и влияния приливов. В глубоких эстуариях фотосинтетическая активность придонной флоры ограниченна, в результате чего суточный ход кислорода выражен довольно слабо. В менее глубоких эстуариях суточные колебания концентрации проявляются ярче. Приливные перемещения воды маскируют суточные колебания и в значительной мере затрудняют их изучение.
Сезонные колебания концентрации кислорода зависят от изменений расхода, приливов, продолжительности дня и биологических процессов. Приток морской воды во время прилива изменяет концентрацию кислорода в эстуарии. Продолжительность дня в значительной степени определяет температуру воды, кроме того, с ней связано функционирование биологических организмов и фотосинтетическая деятельность. Сезонные колебания содержания кислорода хорошо просматриваются на примере Чесапикского залива. В апреле на поверхности содержание кислорода составило 143% равновесного насыщения, а в августе — 42%. В придонных водах концентрация кислорода достигала 133% в октябре и 24% в июне (Reid, 1961).
Значение кислорода в жизни водных организмов
Кислород часто является лимитирующим фактором в водных системах. Источники первичной продукции (фотосинтезирующие растения) при свете продуцируют кислород, но в темноте потребляют его. Таким образом, кислород может лимитировать как первичную, так и вторичную продукцию. Весьма высокое содержание кислорода в атмосфере (около 21%) может замедлять фотосинтез некоторых растений суши. Это явление невозможно в водной среде, так как концентрация кислорода здесь слишком мала. Истощение запасов кислорода в темное время суток может косвенно влиять на фотосинтез, поскольку популяция фотосинтезирующих водорослей сокращается.
Водные организмы исключительно хорошо приспособились к низкому содержанию кислорода и извлекают его из воды даже при очень низких концентрациях. Величина лимитирующей концентрации для любой рыбы зависит от ее генетического строения температуры воды, уровня активности, а также испытываемого стресса. Обычно рыбы не способны существовать при концентрации менее 3 мг/л, хотя некоторые моллюски и другие организмы могут ограниченное время существовать в воде при столь низком содержании кислорода. При концентрации кислорода 3—5 мг/л некоторые виды рыб могут жить неограниченное время, а другие — только в течение короткого периода. При концентрации свыше 5 мг/л все водные организмы могут существовать неограниченное время (в пределах естественного жизненного цикла), если другие параметры окружающей среды благоприятны.
Как правило, низкие концентрации кислорода легче переносятся взрослыми особями видов, стоящих на нижней ступени эволюционной лестницы (например, крабы и устрицы). Молодь почти всех видов менее устойчива к низким концентрациям, чем взрослые. Однако делать широкое обобщение относительно влияния генетического строения и возраста на толерантность к низким концентрациям кислорода трудно из-за значительного разнообразия видов и существующих между ними различий. Потребности в кислороде многих распространенных водных организмов в настоящее время еще неизвестны.
Огромное влияние на потребности в кислороде всех .водных организмов оказывает температура воды. За исключением млекопитающих, большинство водных организмов пойкилотермны, т. е. температура их тела равна температуре окружающей среды. Существует, однако, группа водных организмов, собственная температура которых на несколько градусов выше окружающей. Многие быстроплавающие рыбы, которые постоянно находятся в движении, обладают этой способностью (например, тунцы). При понижении температуры воды происходит соответствующее снижение температуры тела. Метаболическая активность, в сущности, представляет собой цепь ферментативных реакций. При понижении температуры скорость этих реакций уменьшается. Таким образом, интенсивность обмена веществ и активность рыб уменьшаются с понижением температуры, при этом соответственно уменьшается и их потребность в кислороде.
Как уже отмечалось в главе 3, существуют оптимальная и максимальная температуры жизнедеятельности рыб. При оптимальной температуре потребление кислорода высоко в связи с быстрым ростом и значительной активностью. Однако при температуре выше оптимальной организм испытывает стресс, который мобилизует сигнальную и защитную системы организма, что приводит к очень высокому потреблению кислорода. Дальнейшее повышение температуры усугубляет стресс и потребность в кислороде начинает обгонять рост температуры. При испуге и в других стрессовых ситуациях происходит аналогичное увеличение потребности организма в кислороде независимо от температуры. Стресс является одной из причин того, что рыба часто задыхается при вылове.
На потребление кислорода влияет также уровень активности (рис. 4.5). Скорость потребления кислорода непосредственно зависит от его концентрации в воде. Если концентрация столь низка, что организм не может извлечь достаточного для удовлетворения основных метаболических потребностей количества кислорода, наступает гибель. В некотором диапазоне концентраций выше той, при которой уже удовлетворяются основные метаболические потребности, организм может выжить, но не проявлять достаточной активности. При этом он более уязвим для хищников, подвержен заболеваниям и влиянию различных негативных факторов. Поскольку в этой области концентраций рыба находится в стрессовом состоянии, потребности в кислороде возрастают, что еще больше осложняет ее существование. Организмы, способные совершать значительные перемещения, например пелагические рыбы, стремятся по возможности избегать районов, где концентрация кислорода ограничивает их активность. Обычно при недостатке кислорода в поведении рыбы появляется пассивность, она поднимается к поверхности, чтобы глотнуть воздуха. Если персонал, обслуживающий рыбоводные пруды, особенно с плотной посадкой, не обратит внимания на эти особенности поведения рыб, то в течение нескольких минут или часов может произойти катастрофический замор.
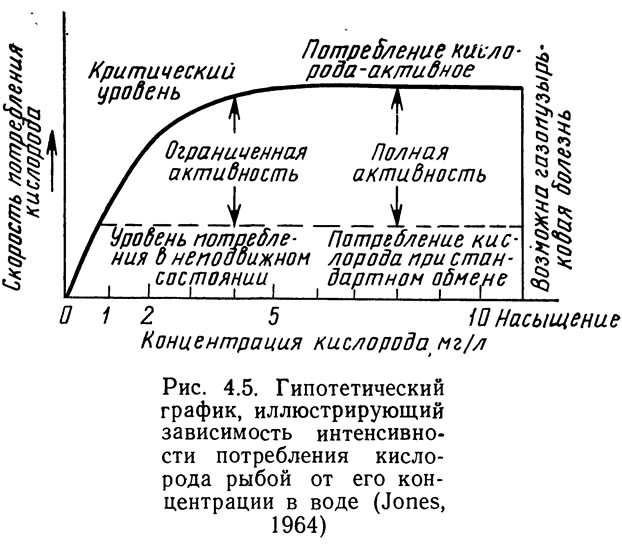
Выше той концентрации, при которой активность организма ограниченна, расположена область концентраций, обеспечивающих полную, неограниченную активность. Нижней границей этой области является критический уровень, показанный на рис. 4.5, а верхней — концентрация равновесного насыщения. Для водных организмов необходимо поддерживать концентрацию кислорода в этом диапазоне. При перенасыщении воды кислородом существует значительный риск развития так называемой газопузырьковой болезни. Она вызывается перенасыщением крови рыб и моллюсков газом, который затем выделяется в виде пузырьков в тканях животного. Накопление большого количества этих пузырьков может привести к гибели. Чаще всего газопузырьковая болезнь вызывается азотом, но кислород также может быть ее причиной.
Необходимо отметить, что критический и летальный уровни, показанные на рис. 4.5, для различных биологических видов могут смещаться как влево, так и вправо. Верхняя граница — уровень равновесного насыщения — определяется исключительно физическими факторами: температурой и соленостью воды. С увеличением температуры воды скорость роста водных организмов возрастает до определенного предела. Температура, обеспечивающая максимальную скорость роста, различна для разных видов (см. главу 3). С увеличением температуры у всех водных организмов, по крайней мере пойкилотермных, потребление кислорода возрастает. Следовательно, повышается и критический уровень концентрации кислорода (см. рис. 4.5). В то же время увеличение температуры приводит к уменьшению концентрации равновесного насыщения. В результате диапазон концентрации кислорода, при котором возможна полная активность, сужается. Это означает, что в системах для культивирования повышение температуры воды вплоть до предельной позволяет ускорять рост культивируемых объектов, но одновременно уменьшает полезный диапазон концентраций кислорода. При этом повышаются требования к контролю концентрации кислорода, особенно в системах с высокой плотностью посадки.
В табл. 4.2 приведены критические концентрации кислорода для нескольких видов рыб при разной температуре. В табл. 4.3—4.5 представлены минимальные концентрации, переносимые различными видами рыб при разных значениях температуры. Из таблиц видно, что при повышении температуры потребности в кислороде одних и тех же видов увеличиваются. Критические концентрации значительно превышают минимальные. Данные для отдельных видов значительно варьируют. Такая вариабельность может быть связана как с различиями в методиках определения критических и минимальных концентраций, так и с индивидуальными особенностями тестируемых особей. Колебания таких факторов, как температура акклимации и уровень стрессовых воздействий, затрудняют определение минимальных и критических концентраций. Значительные трудности возникают при обобщении данных, полученных разными исследователями, как это сделано в табл. 4.2—4.5.

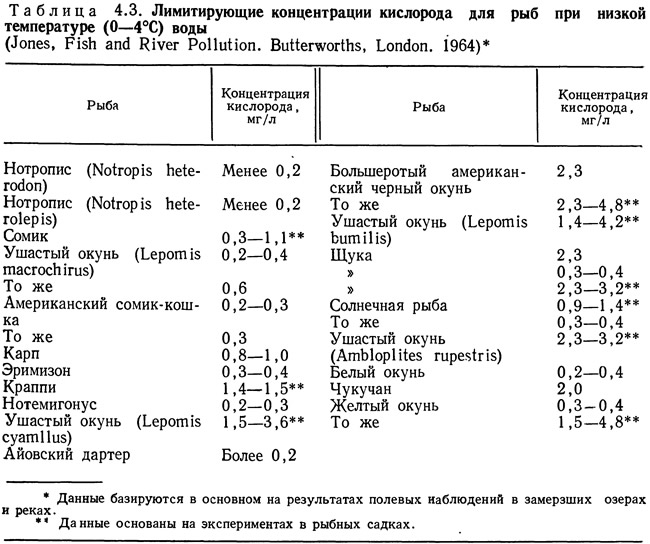

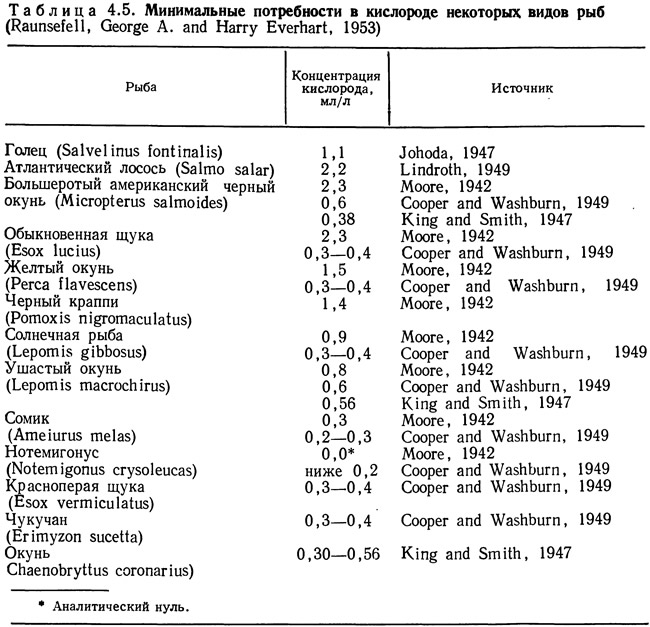
В табл. 4.2—4.5 приведены диапазоны значений или значения критических и минимальных концентраций. Эти данные полезны для конструкторов, но при разработке проектов желательно иметь более полную информацию. Летальные температуры часто выражают с помощью LD50 (температура, при которой 50% подопытных организмов погибают за определенный период времени). Эти сведения также могут быть весьма полезны при проектировании. Однако ни один из этих параметров не учитывает естественных биологических различий между особями. В какой-то мере приближается к этому требованию LD50, но вряд ли найдется инженер, который при разработке проекта будет исходить из 50%-ной смертности культивируемых объектов, поэтому конструкторам необходим показатель, дающий информацию о критических и минимальных значениях параметров с учетом функции распределения. Эта функция должна отражать колебания критических и минимальных температур для отдельных особей при различном времени воздействия. Получение такой информации требует затрат времени и средств, однако она позволяет правильно выбрать кислородный режим в системе для культивирования.
Список литературы
- Cooper, Gerald P. and George N. Washburn (1949). Relation of Dissolved Oxygen to Winter Mortality of Fish in Michigan Lakes. Transactions of the American Fisheries Society 76: 23—33.
- Jahoda, W. Y. (1947). Survival of Brook Trout in Water of Low Oxygen Content. Journal of Wilderness Management 11 (1): 96—97.
- Jones, J. R. Erichsen (1964). Fish and River Pollutioh. Butterworths, London (Reprinted 1966).
- King, Joseph E. and Arthur L., Smith, Jr. (1947). Investigations of the Effect of Water Hyacintn on the Fish and Fish Habitats of Louisiana Waters: U. S. Fish and Wildlife Service, Special Scientific Report 39, App. A: 30—65.
- Lindroth Arne (1949). Vitality of Salmon Parr at Low Oxygen Pressure. Institute of Freshwater Resources, Fisheries Board, Drottningnonh, Sedtn, 29: 49—50.
- Meteorogical Expedition, 1925—27.
- Moore, Walter G. (1942). Field Studies on the Oxygen Requirements of Certain Freshwater Fishes. Ecology 23: 319—329.
- Raymont, John E. G. (1963). Plankton and Productivity in the Oceans. Ma-cmillian, New York.
- Reid, George K. (1961). Ecology of Inland Waters and Estuaries. Reinhold, New York.
- Rounsefell, George A., and W. Harry Everhart (1953). Fishery Science, Its Method and Applications. Wiley, New York.
- Standard Methods for the Examination of Water and Wastewater, 13th ed., 2nd, printing (1971). American Public Health Association, Washington, D. C.
Примечания
1. В отечественной литературе этот слой называется слоем скачка кислорода.
2. С этим утверждением автора трудно согласиться, поскольку колония кораллов не выделяет, а поглощает кислород. В некоторых случаях в воде, прошедшей через коралловый риф, содержится всего 60—70% кислорода.
3. Не являясь специалистом в области химической океанографии, автор ошибочно трактует известный факт о большей аэрируемости промежуточных вод Атлантики в результате меньшей плотностной стратификации.