После достижения максимума при дальнейшем повышении температуры наблюдается уменьшение продукции до наступления гибели. Обычно зависимость величины продукции от температуры описывается уравнением второго или третьего порядка, часто она представляет собой параболу. Такой вид зависимости обычно характерен как для животных, так и для растений.
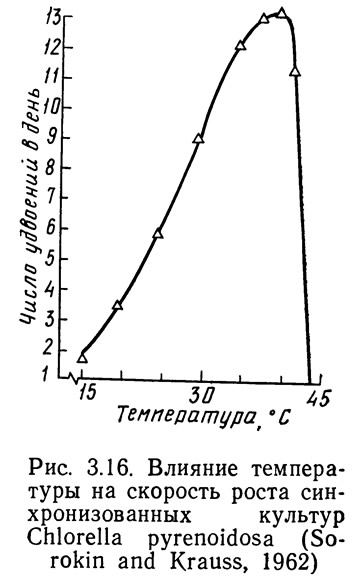
Вид кривой зависимости величины продукции от температуры (см. рис. 3.16) может меняться под влиянием различных факторов. Здесь очень важно определение понятия продукции. Термин «общая продукция» обычно подразумевает продукцию организмов при отсутствии потерь на дыхание. Термин «чистая продукция» используется для определения действительно произведенного, пригодного к потреблению, обычно органического продукта. Общая продукция за вычетом потерь на дыхание равна чистой продукции.
От температуры зависит не только величина общей и чистой продукции, но и дыхание. Как правило, для каждого отдельного организма кривая зависимости дыхания от температуры имеет в основном тот же вид, что и кривая зависимости величины продукции от температуры. Однако, в сущности, это не обязательно, особенно для растений. Если вид кривой дыхания отличается от вида кривой общей продукции, характер кривой величины чистой продукции также должен быть другим.
Влияние температуры на продуктивность любого организма неразрывно связано с воздействием температуры на течение химических реакций. Большинство жизненно важных процессов можно представить как серию химических -реакций, катализируемых ферментами. Обычно при повышении температуры происходит ускорение химических реакций в результате увеличения кинетической энергии атомов. Однако многие соединения, присутствующие в живых организмах, нестабильны при температурах 35—150°С. Некоторые соединения деактивируются при температурах ниже 35°С. Эти факты позволяют объяснить ход кривых зависимости продукции от температуры для большинства организмов. При достаточно низких температурах некоторые принципиально важные для жизнедеятельности реакции не идут вообще или протекают слишком медленно для обеспечения жизнедеятельности организмов. Это и есть минимальная температура, при которой возможно существование организмов. При повышении температуры скорость химических реакций возрастает, что обеспечивает рост организма. При дальнейшем повышении температуры кинетическая энергия становится достаточной для разрушения некоторых необходимых химических соединений. Это уменьшает скорость химических реакций и приводит к снижению скорости роста. При определенных значениях температуры происходит полное разрушение жизненно важных химических соединений, приводящее к гибели организма.
Однако кроме описанных выше изменений химических соединений существуют другие причины гибели организмов, когда температура превышает определенное значение.
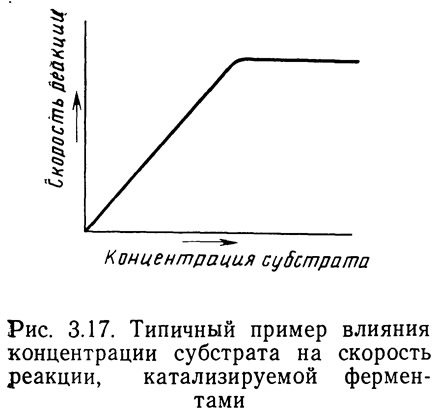
Все или почти все химические реакции, протекающие в живых организмах, катализируются ферментами (органическими катализаторами). В качестве катализаторов они определяют скорость химических реакций. Скорость протекания химических реакций и скорость роста организмов зависят от концентрации ферментов и воздействующей на них температуры окружающей среды. На рис. 3.17 показано влияние концентраций субстрата на скорость реакций. Аналогичный вид имеет график зависимости скорости реакций от концентрации фермента. Увеличение концентрации субстрата приводит к ускорению реакции в результате увеличения числа активных центров. При некоторой концентрации субстрата все ферментное вещество оседает на активные центры. Дальнейшее увеличение числа активных центров при увеличении концентрации субстрата не влияет на скорость реакции, поскольку все ферментное вещество уже использовано, а без него протекание реакции невозможно.
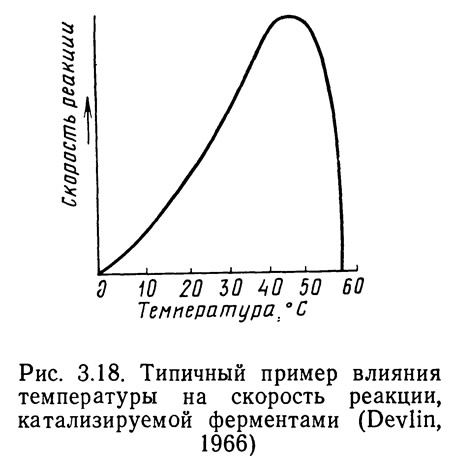
Повышение температуры приводит к увеличению кинетической энергии молекул фермента и субстрата, что вызывает рост числа контактов молекул фермента и субстрата. При этом возрастает скорость реакции, поскольку увеличивается вероятность того, что молекула фермента осядет на активный центр субстрата. Однако при повышении температуры фермент начинает разрушаться, концентрация его уменьшается и реакция замедляется. Каталитические свойства многих ферментов ослабляются при температуре 35°С и полностью утрачиваются при температуре 60°С (Devlin, 1966). На рис. 3.18 показано влияние температуры на скорость катализируемой ферментами реакции. Однако поскольку большинство водных организмов не могут существовать при температурах, превышающих 30—35°С, разрушение ферментов не является основной причиной их гибели. Химические реакции в значительной степени определяют метаболизм пойкилотермных (холоднокровных) животных и именно с этой точки зрения необходимо подходить к обсуждению вопросов термального стресса и гибели животных при повышении температуры окружающей среды.
При проектировании оборудования для культивирования водных организмов необходимо знать температуру, при которой наступает их гибель. Однако этого недостаточно, поскольку характер влияния изменения температуры на рыб необычайно сложен. Необходимо учитывать, по крайней мере, три параметра: температуру акклимации, термального шока и температуру, при которой наступает гибель.
На поверхности жабр кровь рыб практически соприкасается с водой, что облегчает транспорт кислорода, а также интенсивную теплопередачу вследствие высокого коэффициента теплоотдачи воды. Эта теплопередача настолько велика, что рыбы не могут существовать при больших различиях между температурами тела и окружающей водной среды, так как энергетические потребности очень высоки. В процессе эволюции рыбы сформировались как пойкилотермные животные — их ткани способны существовать в некотором температурном интервале; в определенном температурном интервале протекают у них и все биологические процессы. У рыб развилась способность адаптироваться к различным температурам при условии, что температурные изменения происходят не слишком быстро. Адаптация к изменяющейся в определенных пределах температуре окружающей среды называется акклимацией.
Процесс акклимации должен происходить достаточно медленно, чтобы физиологические процессы могли адаптироваться к изменению температуры. При повышении, температуры акклимация происходит значительно быстрее, чем при ее понижении. Для акклимации при повышении температуры на 5°С может потребоваться от нескольких часов до одного-двух дней, тогда как при понижении температуры на такую же величину для этою потребуется несколько дней (Brett, 1956). Замечено, что для некоторых видов рыб низкая концентрация кислорода замедляет акклимацию при повышении температуры (Brett, 1956).
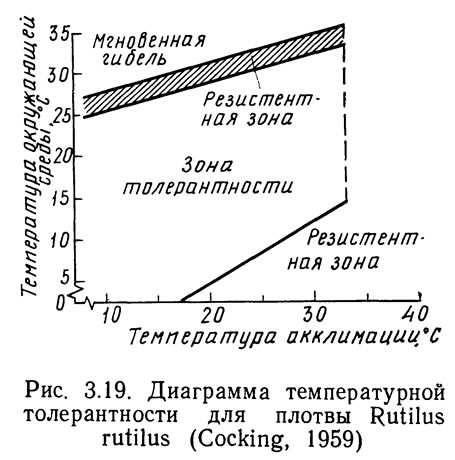
На рис. 3.19 представлена диаграмма температурной толерантности для плотвы. Область толерантности определяет температуру акклимации и окружающей среды, при которых время существования рыб неограниченно. Область резистентности определяет температурный режим, который рыбы могут выдерживать лишь в течение ограниченного периода времени. За пределами области резистентности наступает мгновенная гибель организма. Очевидно, что гибель рыб как в зоне резистентности, так и за ее пределами наступает в результате воздействия высоких температур на их центральную нервную систему, хотя конкретная причина (механизм) такого воздействия неизвестна (Brett, 1956).
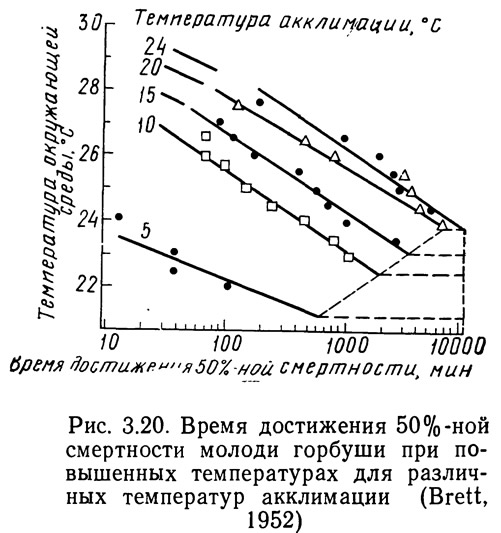
На начальном этапе исследований ученые столкнулись с сложностью определения положения границы между резистентной и толерантной областями, поскольку оно зависит от времени экспозиции. В группе акклимированных рыб, помещенных в испытательную камеру, температура в которой находится в пределах резистентной зоны, наблюдается смертность. Зависимость числа случаев смертности от времени называется кривой смертности. Проводя серию экспериментов, в которых окружающая температура изменяется, а температура акклимации остается постоянной, можно получить серию кривых смертности. Зависимость логарифмов времени достижения 50%-ной смертности от температуры представлена на рис. 3.20. Эта полулогарифмическая зависимость линейна, причем наклон прямых почти одинаков для всех видов, хотя отрезки, отсекаемые ими на оси координат, различны. Существенно, что в определенный момент времени наклон кривой внезапно изменяется и она становится горизонтальной (Brett, 1956). Дальнейшее увеличение времени экспозиции не вызывает возрастания смертности. Температура, при которой происходит изменение наклона, является летальной и соответствует переходу из резистентной зоны в толерантную.
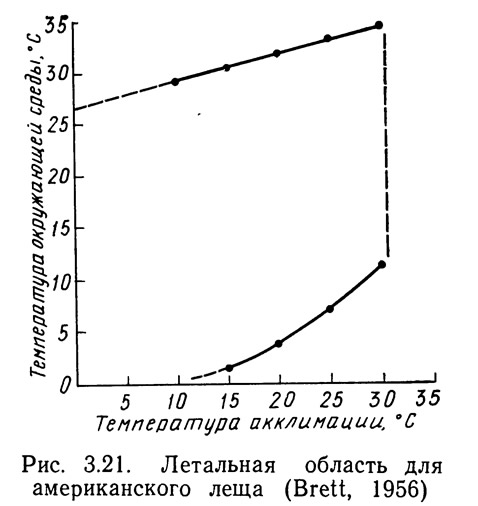
Изменение температуры акклимации приводит к изменению и летальной температуры. Повышение температуры акклимации обычно вызывает увеличение верхней, а иногда также и нижней летальной температуры; понижение температуры акклимации дает противоположный эффект. Данной температуре акклимации и температуре LD50 (температуре, при которой гибнут 50% подопытных животных) соответствует одна точка на границе области в двухмерном пространстве. На рис. 3.21 представлена летальная область для американского леща. Отметим, что при повышении температуры акклимации на 5°С верхняя летальная температура возросла всего на 1,3°С. В данном случае нижняя летальная температура при повышении температуры акклимации изменяется нелинейно, хотя для некоторых рыб характерен ее линейный рост; скорость повышения нижней летальной температуры увеличивается при температурах акклимации, превышающих 12°С.
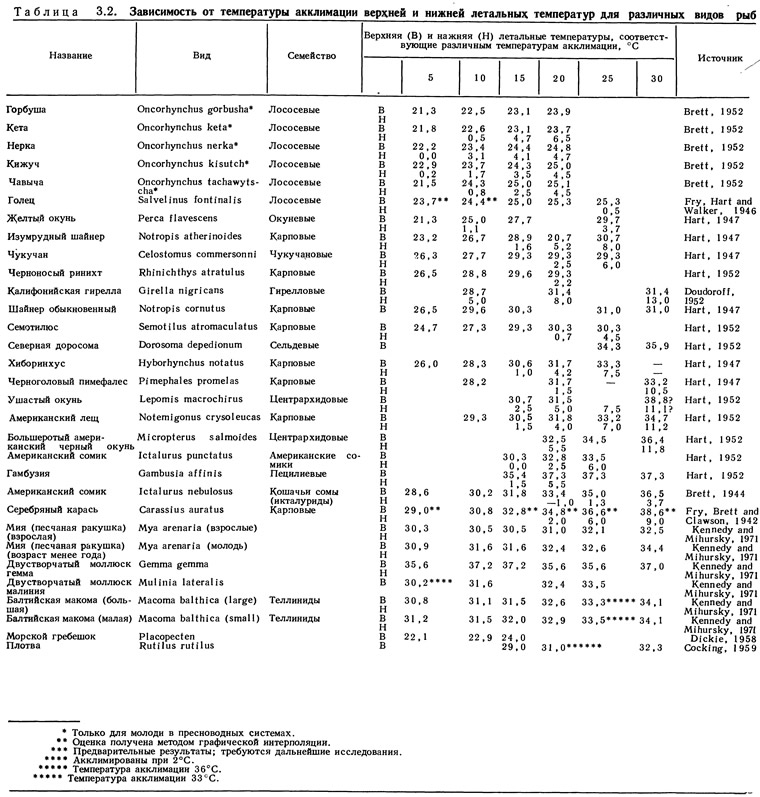
Конфигурация, размеры и другие характеристики летальной области специфичны для различных видов, но вполне воспроизводимы для каждого данного вида. Значение летальной температуры для любого вида рыб при температуре акклимации может быть воспроизведено с точностью до 0,2°С (Brett, 1956). Однако имеющихся данных недостаточно для построения замкнутых летальных областей для всех видов рыб. Значительная часть имеющихся данных сведена в табл. 3.2.
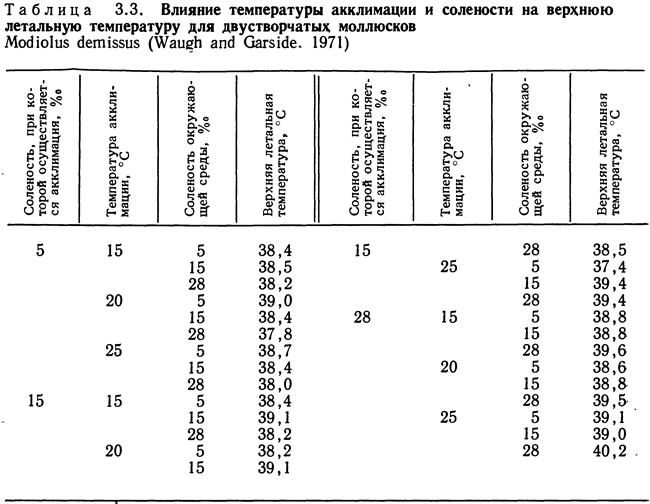
Летальные области строятся для условий, при которых стресс вызывается только температурными изменениями. Обычно при наличии других параметров, вызывающих стресс, значение летальной температуры изменяется. Проводились исследования влияния изменения температуры акклимадии и солености на верхнюю летальную температуру двустворчатых моллюсков Modiolus demissus (Waugh and Garside, 1971) (табл. 3.3). Полученные изменения верхней летальной температуры весьма значимы, хотя и малы по абсолютной величине.
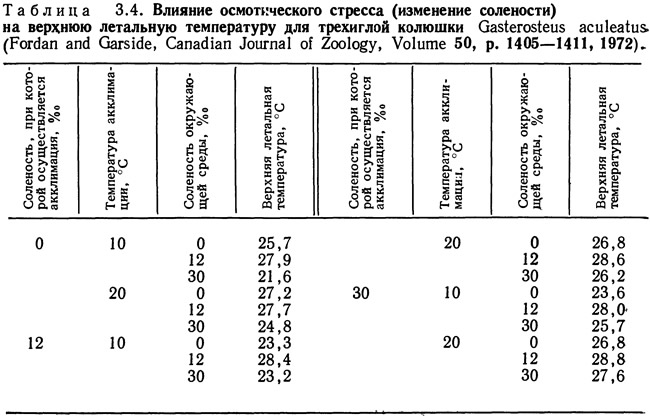
Верхние летальные температуры для фундулюса Fundulus diaphanus составляли 27,5, 33,5 и 27,5°С при соленостях 0, 14 и 32% соответственно (Garside and Jordan, 1968). Аналогичные данные относительно влияния солености на верхнюю летальную температуру для трехиглой колюшки Gasterosteus aculeatus приведены в табл. 3.4. (Jordan and Garside, 1972). Наивысшая летальная температура была зарегистрирована для солености, равной 12‰, независимо от солености и температуры акклимадии. При отклонении солености от этого значения в любую сторону значение верхней летальной температуры уменьшается. Стрессы, вызванные другими факторами, также понижают верхнюю летальную температуру.
Летальная температура является важным фактором, определяющим возможность существования данного вида в определенных природных условиях. Однако уже при температурах несколько ниже температур верхней резистентной зоны, а также при температурах выше соответствующих границе нижней резистентной зоне рыбы находятся в состоянии стресса и теряют свою конкурентоспособность, поскольку они не могут нормально функционировать. При создании искусственных систем обычно стремятся к получению максимальной прибыли. Поскольку содержание рыб, находящихся в состоянии стресса, менее выгодно, температурный режим системы не должен устанавливаться исходя из значений летальных температур. Допустимое сужение температурного интервала определяется из условий минимизации стресса.
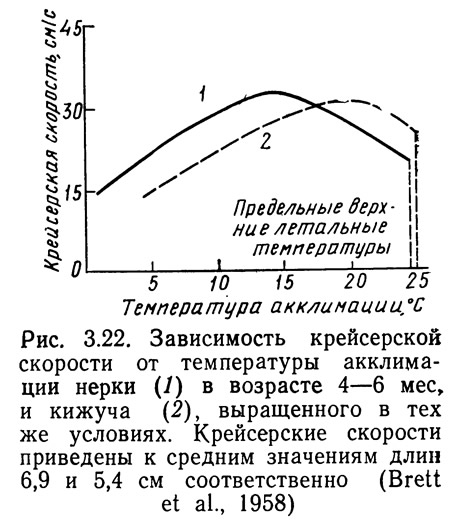
Для определения степени температурного стресса рыб обычно используют косвенные методы измерения. Один из них заключается в измерении крейсерской скорости рыб при различных температурах. На рис. 3.22 представлена зависимость крейсерской скорости от температуры акклимадии: при увеличении температуры скорость сначала достигает максимума, а затем убывает. Температура, соответствующая наибольшей скорости, обеспечивает оптимальные условия выживания рыб. Она несколько ниже минимальной температуры термального стресса.
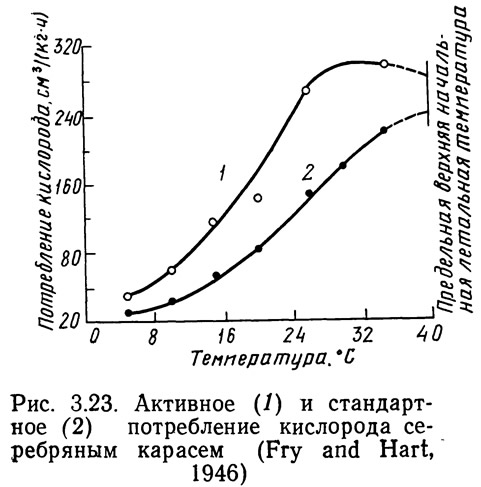
Косвенным методом определения влияния термального стресса может служить измерение скорости метаболизма или такого его показателя, как потребление кислорода. Фрай и Харт (Fry and, Hart, 1946) определяли зависимость стандартного и максимального потребления кислорода от температуры для серебряного карася Carassius auratus. Стандартное потребление кислорода — количество кислорода, необходимое для поддержания основного обмена при определенной температуре. Кривая максимального продолжительного потребления кислорода строится с учетом стандартного потребления кислорода, кислорода, необходимого для активной жизнедеятельности, а также поглощенного сверх основного уровня обмена при увеличении интенсивности дыхания и кровообращения. Если последней величиной пренебречь или объединить ее с потреблением кислорода, необходимого для поддержания жизнедеятельности, разность между максимальным и стандартным потреблением кислорода даст величину потребления кислорода, необходимого для поддержания жизнедеятельности при любой температуре. На рис. 3.23 приведены скорости максимального (активного) и стандартного потребления кислорода в зависимости от температуры. На рисунке 3.24 показаны кривые активного и стандартного потребления кислорода. Обе скорости возрастают при повышении температуры до определенной величины.
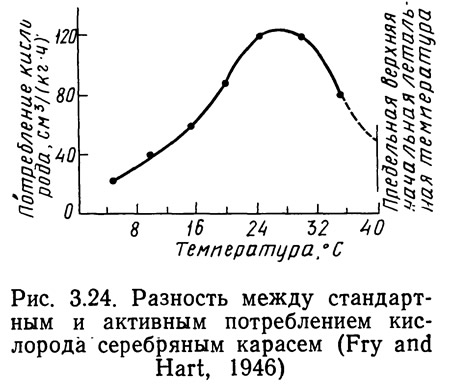
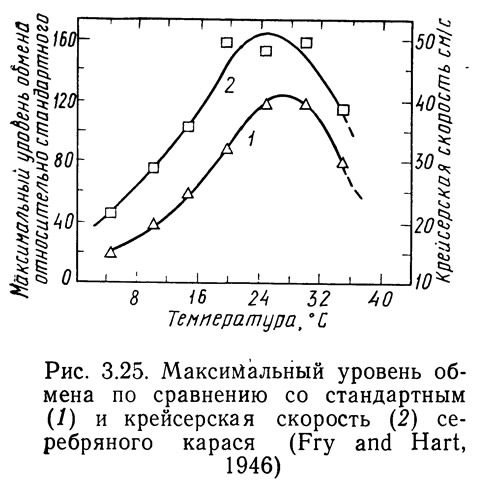
При повышении температуры активное потребление кислорода снижается быстрее, чем при стандартном обмене. Из рис. 3.24 видно, что активное потребление кислорода серебряным карасем возрастает при повышении температуры до 28°С, а затем быстро снижается. Область температур выше 28°С для серебряного карася — область термального стресса (Fry and Hart, 1946). Фрай и Харт сопоставляли также зависимости максимальной крейсерской скорости и скорости потребления кислорода для серебряного карася от температуры (рис. 3.25). Кривые имеют почти одинаковую форму, что доказывает сопоставимость результатов, полученных различными косвенными методами, и применимость этих методов для определения момента наступления температурного стресса.
Оба описанных метода полезны для оценки условий внешней среды, так как измеряемые параметры влияют на выживаемость организма. Поскольку в искусственных промышленных системах основная доля затрат приходится на приобретение корма для рыб, показателем эффективности системы является эффективность утилизации пищи и (или) скорость роста рыб.
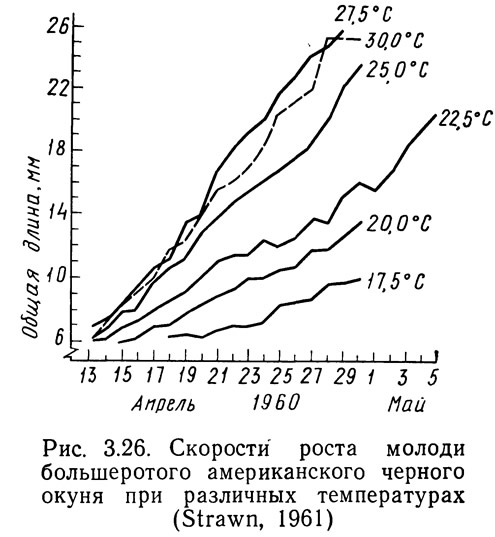
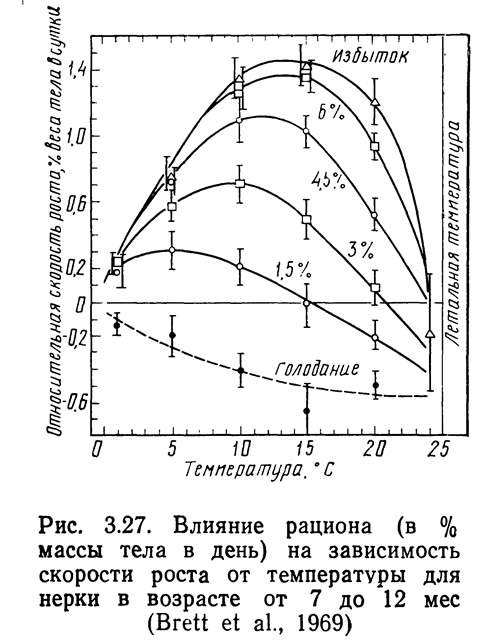
Простой метод сравнения эффективности, основанный только на определении изменения скорости роста в зависимости от времени и температуры, представлен на рис. 3.26. Повышение температуры до 27,5°С вызывает увеличение скорости роста молоди большеротого окуня. Этот метод можно использовать для естественных- водоемов, в которых невозможно контролировать количества пищи и другие факторы, влияющие на рост рыб. Если же корма приходится закупать, удобнее применять метод анализа, аналогичный разработанному Бреттом (Brett et al., 1969). На рис. 3.27 показано влияние температуры на скорость роста нерки при различных режимах питания. Оптимальный рост наблюдался при избыточном питании, причем при увеличении количества потребляемой пищи температура, обеспечивающая максимальный рост, повышалась, при высоких температурах рыбы голодали даже тогда, когда количество потребляемой пищи составляло 3% веса тела в день. Когда скорость метаболизма является функцией температуры, нет ничего удивительного в том, что при высоких температурах рыбы голодают даже при наличии корма. Метаболические характеристики также позволяют объяснить повышение температуры, соответствующей максимальной скорости роста, при увеличении рациона.
При эксплуатации хозяйств аквакультуры интерес представляет не столько скорость роста, сколько получаемая прибыль. Рис. 3.28 получен наложением кривых эффективности на кривые скорости роста, приведенные на рис. 3.27. Кривые общей эффективности были рассчитаны по следующему уравнению:
где Eg — эффективность утилизации пищи; G — прирост; I — потребление пищи.
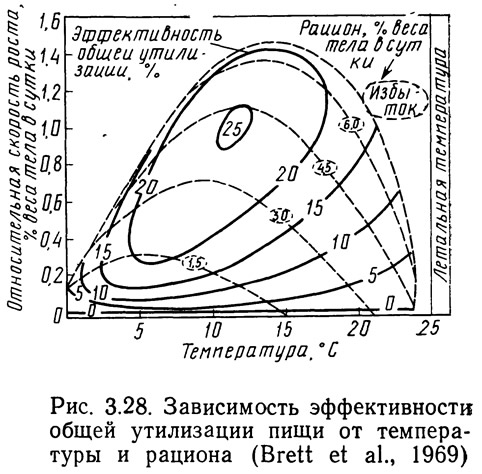
Эффективность утилизации пищи может изменяться от 0 до 25%. Для получения максимальной прибыли при учете только затрат на корм нерку необходимо содержать в диапазоне температур 10—13°С и стремиться к приросту около 1% веса.
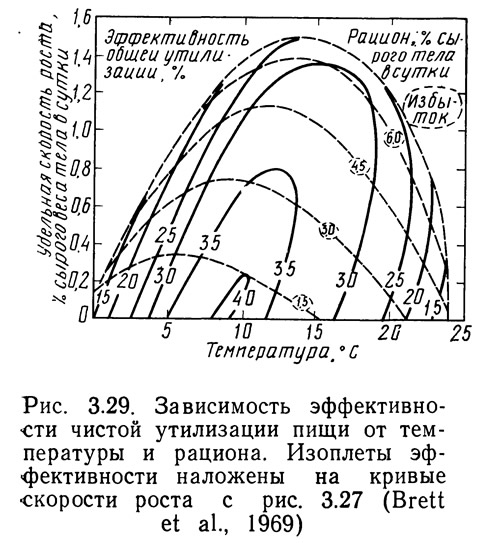
Эффективность общей утилизации не является важным параметром с рыбоводной точки зрения. Эффективность чистой утилизации позволяет точнее оценить вес рыбной продукции, полученной на единицу веса корма. Чистая утилизация определяется следующим образом:
чистая утилизация пищи = общая утилизация пищи — затраты на обмен. (3.4)
На рис. 3.29 приведены зависимости скорости роста от температуры и рациона; на график нанесены также кривые чистой утилизации пищи. Такой график представляет больший интерес для рыбовода, чем график типа, показанного на рис. 3.28.
При искусственном разведении рыб кроме затрат на корма существуют и других расходы, которые необходимо учитывать при оценке эффективности системы. Существует несколько методов учета этих затрат, одним из которых является метод многомерного .анализа, аналогичный проиллюстрированному рис. 3.28, но с включением всех затрат. Очевидно, что ввиду его многомерности полученное уравнение не может быть представлено на одном графике, однако для нахождения оптимального решения может быть проведена максимизация математических уравнений. Такой анализ желательно проводить всегда, но он требует большого количества экспериментальных данных. Даже данные, аналогичные приведенным на рис. 3.28, собраны только для некоторых видов рыб, таких: как нерка и сомик. Однако и для этих видов данных недостаточно для проведения упомянутого выше многомерного анализа.
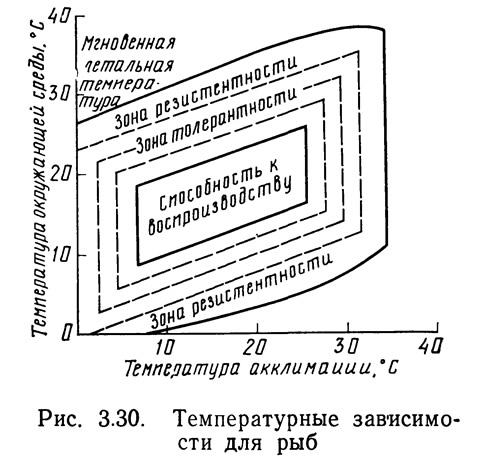
Полученные для рыб температурные зависимости представлены на рис. 3.30. Внешняя линия определяет максимальные значения температур, при которых может существовать рыба. Следующая линия (первая штриховая) показывает, при каких максимальных значениях температур рыба может существовать неограниченное время. Эта линия служит границей между зонами резистентности и толерантности. Внутри зоны толерантности существует несколько уровней термального стресса, когда существование рыбы возможно, хотя ее активность, скорость роста или эффективность утилизации пищи снижаются. Внутренний четырехугольник определяет термальные границы возможного воспроизводства, т. е. значения температур, не вызывающих или почти не вызывающих стресса организмов. Термальный стресс в областях, прилегающих к зоне воспроизводства, может быть выявлен далеко не всегда. К сожалению, ни для одного из видов не собрана информация, необходимая для полного построения диаграммы, приведенной на рис. 3.30; для большинства видов данные отсутствуют вообще или их недостаточно для построения основных элементов диаграммы.