Содержащиеся в кормах жиры оказывают влияние на их химические свойства и структуру, включая прочность и водостойкость, на вкус и аромат, а также привлекательность для рыб.
Жиры представляют собой сложную смесь органических соединений, различных по своей химической природе, но обладающих общим специфическим свойством — нерастворимостью в воде и растворимостью в органических растворителях (этаноле, ацетоне, эфире, гексане, хлороформе и т.д.). По принятой в зоотехнии схеме химических анализов эти соединения объединяются в группу "сырой жир".
По химическому составу жиры относятся к классу эфиров и делятся на простые (липиды) и сложные (липоиды).
Простые липиды — это эфиры трехатомного спирта глицерина (а также ряда других высокомолекулярных спиртов) и разнообразных жирных кислот. В зависимости от того, сколько гидроксильных групп глицерина замещено жирными кислотами, они носят название моно-, ди- и триацилглицеринов. К этой группе относятся еще воски и стериды.
Сложную смесь триглицеридов (или триацилглицеринов) обычно называют нейтральными липидами, нейтральными жирами или просто жирами [Кизивегтер, 1973].
Триглицериды — главный энергетический резерв организма. Они концентрируются в подкожной жировой ткани, тканях, окружающих внутренние органы — кишечнике, гонадах, селезенке, костях, а также в органах, активно участвующих в жизнедеятельности организма — печени, мозге, мышцах. В клетках тканей они очень тонко диспергированы, вследствие чего на большой поверхности соприкасаются со многими веществами, входящими в состав клеточных структур. Будучи плохими проводниками тепла, они предохраняют внутренние органы от охлаждения, участвуя в процессах теплорегуляции. Кроме того, упругая жировая прослойка, облегающая отдельные органы, одновременно играет роль амортизатора. В качестве запасных липидов они являются подвижной составной частью тканей, и уровень их накопления находится в прямой зависимости от упитанности.
В одноименных тканях различных видов рыб липиды накапливаются избирательно. Их содержание в мышцах находится в пределах 0,2-28%, печени — 0,5-80%, костной ткани — 0,5-2%, крови, слизи — 0,01-0,05%, жировой ткани — 42-92% [Кизиветтер, 1973].
Липоиды, или сложные липиды. Многочисленная группа жироподобных веществ. Среди них очень важную физиологическую роль играют фосфолипиды.
Фосфолипиды (или фосфатиды) представлены многообразной группой веществ различного состава и структуры. Это сложные эфиры многоатомных спиртов и высокомолекулярных жирных кислот, соединенных с фосфорной кислотой и азотистыми основаниями. Радикалы фосфорной кислоты несут отрицательный заряд, поэтому группа имеет еще одно название — полярные липиды. В число наиболее часто встречающихся азотистых соединений входят холин, этаноламин, серии.
К фосфолипидам, образованным на основе глицерина, относят лецитины (например, фосфатидилхолин, лизолецитин), кефалины, серинфосфатиды, а на основе других спиртов (сфингозина, инозитола) — сфингофосфатиды и инозитфосфатиды.
Фосфатиды в виде протеолипидных комплексов входят в состав оболочек и протоплазмы клеток всех животных организмов. Вместе с другими липидами они образуют периферическую мембрану клеток и их органелл.
Функциональное значение фосфолипидов обусловлено их бифазным характером, который позволяет регулировать проницаемость клеточных оболочек. Одна часть молекулы, содержащая жирные кислоты, способна пропускать через себя жирорастворимые вещества, другая — водорастворимые. Фосфолипиды поддерживают работу таких клеточных механизмов, как ионный обмен, дыхание, биологическое окисление. Они влияют на процессы окислительного фосфорилирования в митохондриях, где их содержание достигает 30%. Из-за недостатка в организме рыб фосфолипидов нарушается жировой обмен, что вызывает жировое перерождение печени, нередко приводящее к патологическим изменениям этого органа. В составе мозговой ткани фосфолипиды (в частности лецитины) обеспечивают нормальное функционирование нервной системы. Поддерживая постоянство внутренней среды и межклеточный обмен, фосфолипиды способствуют адаптации организма к изменяющимся условиям. В общих липидах рыб фосфолипиды составляют 10-20%, а иногда и более.
Содержание структурных и плазматических липидов отличается относительным постоянством и, практически, не зависит от упитанности или истощенности организма. Состав запасных липидов определяется в основном особенностями пищевых жиров.
Жирные кислоты. Химические и физиологические свойства различных липидов во многом зависят от типа и количественного содержания жирных кислот. По характеру химических связей их делят на насыщенные, в которых все атомы углерода соединены между собой одной связью, и ненасыщенные, обычно имеющие от 1 до 6 двойных связей и получившие название моно- и полиненасьнценных (или моно- и полиеновых). По положению первой двойной связи (по отношению к последнему атому углерода в концевой метильной группе, что обозначается символом п или (о) ненасыщенные жирные кислоты делятся на следующие семейства (или ряды): линолената (п-3), линолеата (п-6), пальмитоолеата (п-7) и олеата (п-9). При увеличении числа углеродных атомов и двойных связей в жирных кислотах температура их плавления понижается (рис. 1).
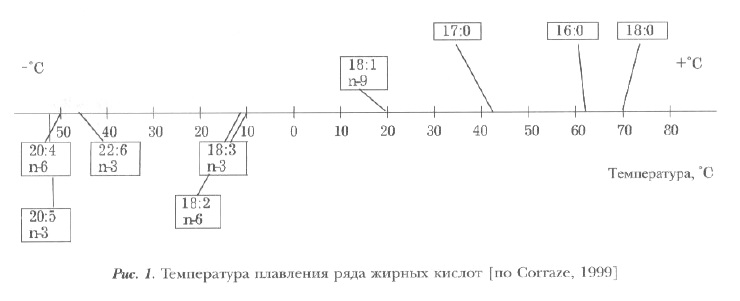
В липидах растений преобладают ненасыщенные кислоты, поэтому растительные масла имеют жидкую консистенцию и низкую точку плавления. В жирах теплокровных животных преобладают насыщенные и мононенасыщенные кислоты (олеиновая, пальмитиновая, стеариновая). Поэтому их жир имеет более плотную консистенцию и более высокую точку плавления. Их основные полиненасыщенные кислоты — линолевая и арахидоновая, принадлежат к семейству линолеата (п-6).
Липиды теплокровных животных при правильном питании обычно имеют такой состав жирных кислот, который при температуре тела позволяет организму поддерживать жидкую консистенцию жира. Для липидов рыб как пойкилотерм-ных животных характерны значительно более разнообразные жирные кислоты с большим числом углеродных атомов (12-24). Среди них преобладают ненасыщенные жирные кислоты — их около 70%, в том числе с 1-6 двойными связями.
По сравнению с теплокровными животными у рыб в составе полиненасыгцен-ных жирных кислот заметно больше кислот ряда линолената (п-З). Соотношение полиеновых кислот п-З/п-6 рядов служит отличительным признаком липидов морских и пресноводных рыб. У морских рыб оно значительно шире, чем у пресноводных (известные диапазоны составляют соответственно 5-14 и 1-4). Причем в липидах морских рыб преобладают эйкозопентаеновая (20:5 п-З) и докозогексаеновая (22:6 п-З) кислоты. У пресноводных холодноводных рыб доля п-З кислот с 5 и 6 двойными связями также выше, чем у тепловодных. Это создает необходимую жидкостность структуры клеточных мембран и позволяет поддерживать их нормальную проницаемость при более низких температурах, чем у тепловодных рыб, обитающих в пресной воде.
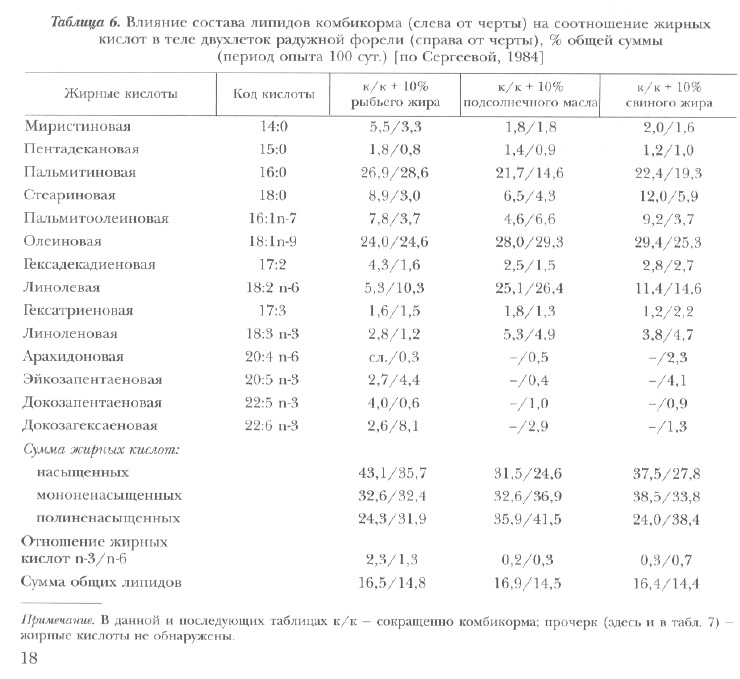
Одним из факторов, от которых зависят липидный состав тканей рыб, их вкус и сроки хранения рыбной продукции, является состав липидов пищи (табл. 6, 7). Так, при длительном питании радужной форели кормом с преобладанием тех или иных жирных кислот изменяется их содержание в общих липидах. В случае обогащения корма рыбьим жиром наиболее существенные сдвиги отмечены в содержании докозагексаеновой, пальмитиновой (увеличение), а также линолевой, линоленовой и арахидоновой (снижение) кислот. При использовании масла подсолнечника в теле рыб отмечено повышение уровня линолевой и линоленовой кислот и понижение — эйкозопентаеновой и докозагексаеновой. В варианте со свиным жиром возрастает доля стеариновой и арахидоновой кислот, сокращается — линолевой и докозагексаеновой.
Сходство состава липидов пищи и организма рыб обусловлено существованием двух основных путей всасывания жира в пищеварительном тракте: первый — после полного предварительного расщепления (липолиза), второй — без него. При липолизе жир распадается на глицерин и жирные кислоты. Жирные кислоты с короткой углеродной цепью активным путем транспортируются через слизистую оболочку кишечника в кровь и далее поступают в воротную вену и печень. Здесь в цитоплазме рабочих клеток печени — гепатоцитах — они используются для синтеза тканевых липидов или текущих метаболических потребностей. Кислоты с длинными углеродными цепями, превращаясь в триглицериды, вместе с холестерином и фосфолипидами образуют микроскопические капельки, заключенные в белковые глобулиновые оболочки, так называемые "хиломикроны".
Последние переходят в лимфу, а затем в кровь, переносящую их к печени. В тканях печени жирные кислоты либо используются для синтеза собственных структурных липидов, либо вновь поступают в кровь и разносятся по организму, где могут депонироваться в форме триглицеридов. Фосфолипиды подвергаются ферментативному расщеплению на составные части. Всасываясь в кишечнике, продукты их распада поступают в печень, где из них синтезируются фосфолипиды, специфичные для данного организма.
Обмен жирных кислот в жировой ткани зависит от степени обеспеченности животного организма энергией. При избыточном поступлении энергии с кормами жирные кислоты липидов корма остаются в жировой ткани. При низком уровне жирных кислот в рационе удовлетворение потребности организма идет за счет депонированных жирных кислот.
Окисление жирных кислот, происходящее в митохондриях клеток, является одним из наиболее эффективных путей снабжения организма энергией. При окислительном распаде 1 г жира организм рыб получает 39,4 кДж энергии, тогда как при распаде 1 г белков — 22,9 и 1 г углеводов — 16,4 кДж. Распадаясь в организме, жиры дают не только энергию, но и воду. При окислении 1 г жиров выделяется 1,07 г воды, 1 г белка — 0,41 г, 1 г углеводов — 0,55 г.
Помимо пищи, источниками жирных кислот служат углеродные остатки углеводов, белков и жирных кислот тканей. Из 100 г белка может образоваться 51 г, а из 100 г крахмала — 41 г липидов.
Однако часть полиненасыщенных жирных кислот не может синтезироваться в организме и должна поступать с пищей. Поэтому их, также как аминокислоты, относят к незаменимым факторам питания.
Биологическая активность эссенциальных кислот определяется местом расположения двойных связей и пространственной конфигурацией этих двойных связей в молекуле. Наибольшей активностью обладают цис-изомеры, у которых первая двойная связь находится между 3-м и 4-м атомами углерода от концевой метальной группы. Поэтому в число незаменимых жирных кислот входит линоленовая с тремя двойными связями (18:3 п-3). Высоко активна и линолевая кислота, имеющая две двойные связи (18:2 п-6).
Кроме них, физиологически важными по выполняемым функциям для рыб считаются их производные: арахидоновая кислота (20:4 п-6) с четырьмя, эйкозопентаеновая (20:5 п-З) и докозапентаеновая (22:5 п-З) с пятью и докозагексаеновая (22:6 п-З) с шестью двойными связями.
Последние у некоторых видов рыб могут образовываться из линолевой и линоленовой кислот путем ферментативной элонгации (удлинения цепи) и десатурации (образования двойных связей).
Наиболее типичные признаки дефицита незаменимых жирных кислот — это снижение аппетита, замедление роста, заболевания кожи и плавников (выражаются в нарушении пигментации и последующем некрозе), повреждение хлоридных клеток жабр, осуществляющих осморегуляцию. Нарушения липидного обмена проявляются в повышенном отложении жира в печени и на внутренних органах. Нарушается воспроизводительная функция, снижается иммунитет. В результате повышается восприимчивость рыб к инфекциям и т.д. Большинство нарушений наблюдается у ранней молоди или производителей до и после нереста, при голодании или недостаточно полноценном питании.
Обобщенные данные о качественных и количественных потребностях рыб в жирных кислотах приведены в табл. 8.
Число незаменимых жирных кислот и потребность в них зависят от видовой принадлежности рыб, температурных условий их обитания и солености воды. Пресноводные холодноводные рыбы в основном нуждаются в п-З кислотах линоленового ряда: линоленовой (18:3 п-З), эйкозапентаеновой (20:5 п-З) и докозагексаеновой (22:6 п-З). В то же время форель способна путем десатурации легко преобразовывать линоленовую кислоту (18:3 п-З) в кислоту 20:5 п-З и 22:6 п-З. Однако по некоторым данным [Takeuchi, Watanabe, 1977], молодь холодноводных рыб обладает более ограниченной способностью к элонгации и десатурации, т.е. синтезу высоконенасыщенных жирных кислот с большим числом углеродных атомов, и поэтому ее потребности в пента- и докозагексаенах выше, чем у взрослых. Отмечено, что способность у форели к биоконверсии жирных кислот существенно выше, чем у морских рыб. Если принять ее за 100%, то у аю она составит 36, угря — 20, дорады — 15, мраморного морского окуня — 7% [Corraze, 1994]. Поэтому морские рыбы больше нуждаются в кислотах 20:5 п-З и 22:6 п-З. Линоленовая кислота в их питании не эффективна.
Для теплолюбивого карпа необходимы линолевая, линоленовая и арахидоновая кислоты. Тиляпия имеет четко выраженную потребность в кислотах линолевого (п-6) ряда и слабо растет при высоких уровнях линоленовой кислоты (п-3 ряд). Для морских рыб требуется высоконенасыщенные кислоты п-З ряда с 5 и 6 двойными связями, т.к. они обладают ограниченной способностью преобразовывать линоленовую кислоту в пентаены и гексаены.
Уровень общих липидов в комбикормах для различных объектов аквакультуры может варьировать в широких пределах в зависимости от биологии и характера питания вида, возраста, условий среды (температуры, солености) и способа выращивания рыб (в прудах, садках, рециркуляционных системах). Оптимальный уровень липидов в кормах для пресноводных (форели, карпа, тиляпии) и морских (лаврака, тюрбо) рыб [по Corraze, 1999] приведен ниже.
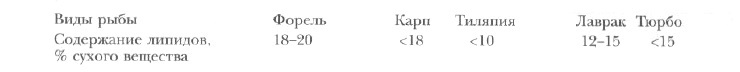
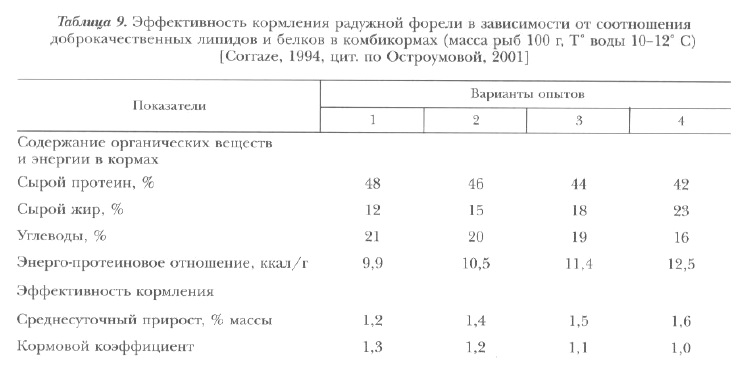
Наивысшая эффективность использования корма у радужной форели получена при содержании белка и жира в рационе соответственно 44 и 18%, а также 42 и 23% (табл. 9).
Карп без видимых вредных последствий может переносить до 40% доброкачественного жира в диете [Kaushik, 1995]. Нижняя граница 3-2,5%. При ее снижении нарушается нормальный ход обменных процессов, что приводит к снижению эффективности использования корма и, в частности, белка. В этом случае организм вынужденно использует в процессах пищеварения эндогенные запасы липидов и часть их удаляется с экскрементами [Щербина, 1973а].
Известно, что жиры как источник энергии обладают "белоксберегающим" действием. В японской практике для индустриального карпа при увеличении в комбикормах доброкачественных жиров до 18% количество белка снижали с 45 до 29% без ухудшения продукционного эффекта, у форели сходный эффект наблюдали при увеличении жира с 15 до 20% и соответственно снижении протеина с 48 до 35%. Аналогичные исследования имеются по белокорому палтусу. Уровень полноценного жира в корме форели, равный 25-30%, также не вызывает нежелательных отклонений в метаболизме и росте рыб. По имеющимся данным, у атлантического лосося при увеличении уровня жира с 22 до 30% усвоение пищи повышается на 10% и отходы органического вещества, аммония и фосфора снижаются на 23, 35 и 22% соответственно. Уровень жира в корме, равный 33-36%, у лососей, выращиваемых на морских фермах Норвегии, также оказывает положительное действие. Причем лучший прирост массы и оплата корма отмечены у рыб, питавшихся кормом с 36% жира в зимний период (3°C, продолжительность светового дня 6 ч). Летом (при температуре 15°C, продолжительности светового дня 20 ч) лучший эффект дал корм с 33% жира. Сезонные изменения температуры влияют на соотношение между основными питательными веществами корма. По мнению норвежских исследователей, зимой содержание жира в корме лососей должно быть значительно выше, чем с летом, тогда как доля белков должна быть летом выше, чем зимой [Rprvik et al., 1998, цит. по Мирзоевой, 1999].
Промышленные корма для форели в настоящее время выпускаются западными фирмами (например, "Эвос") с содержанием 20-22% жира, 44-52% протеина и до 1% клетчатки. Проблема в технологических приемах сохранения доброкачественности жира, т.е. предотвращении его окисления и порчи.
Большое значение имеет качественный и количественный состав питательных веществ в рационах. Высокий уровень жира в кормах может изменять потребность рыб в других элементах питания. Так, в комбикормах для лососей с 15%-ным уровнем жира для устранения симптомов недостаточности витамина Е потребовалось в 3,3 раза увеличить обычно рекомендуемые дозы а-токоферола (до 100 мг/кг корма). С повышением уровня витамина Е улучшается качество выращиваемой продукции (ее цвет, вкус, увеличиваются сроки хранения [Albrektsen, Wathen, 1997].
Жиры, входящие в состав сырья или жировых добавок, обладают способностью в присутствии кислорода воздуха окисляться по типу перекисного окисления. Активизации процессов окисления способствует фермент липоксидаза, которая присутствует в большинстве видов сырья. В результате комбикорма становятся токсичными. Вредное влияние прогорклых жиров на рыб известно давно. Оно выражается в торможении роста, возникновении анемии, нарушении жирового обмена (уменьшается отложение полиненасыщенных жирных кислот), повреждении мускулатуры и снижении общей резистентности и выживаемости рыб. Известен синдром "усыхающей спины".
Способность липидов к окислению зависит от их состава, технологии изготовления и хранения исходного сырья и комбикормов. Более жирное сырье прогоркает быстрее, чем менее жирное. Жиры, содержащие большее количество полиненасыщенных жирных кислот, окисляются быстрее, чем насыщенные (например, рыбий жир прогоркает быстрее, чем говяжий). Специфический запах рыбьего жира обусловлен продуктами окисления докоза- и эйкозапентаеновых и гексаеновых кислот, запах льняного масла — линоленовой, оливкового — олеиновой и т.д.
Окислению липидов способствуют повышение температуры, влажности, развитие микроорганизмов, разлагающих питательные вещества кормов, в частности белки. В окислении липидов участвуют окислительные ферменты собственно кормов (например, липоксидазы соевого, подсолнечного и других видов жмыхов и шротов), а также ферменты, вырабатываемые развивающейся микрофлорой.
Этот процесс приводит к образованию перекисей в результате реакции свободных и связанных жирных кислот с молекулярным кислородом воздуха. Для его начала необходимы повышенная температура (15-30°C) и присутствие влаги. На свету он усиливается. Образовавшиеся перекиси инициируют свободнорадикальное окисление липидов, белков и углеводов. Накопление продуктов перекисного окисления жира в кормах вызывает разрушение содержащихся в них витаминов А, Д, Е, В, и особенно витамина С, а также незаменимых аминокислот [Ржавская, 1976]. Окисление липидов может усиливаться при наличии в комбикормах больших количеств рыбной муки, которая обычно богата не только высоконенасыщенными жирными кислотами, но также белками и железом.
Все перечисленное приводит к резкому снижению продуктивного действия комбикормов. Поэтому при применении комбикормов, особенно с высоким содержанием рыбной муки или жировых добавок, необходим контроль за соблюдением условий хранения и сроков годности; обязательна лабораторная оценка качества входящего в их состав жира.
Для оценки качества жира наиболее информативным считается такой показатель, как кислотное число, характеризующее степень гидролиза жиров и наличие свободных жирных кислот, которые быстрее, чем триглицерины (нейтральные жиры), подвергаются окислению с образованием вредных веществ. В процессе окисления жиров этот показатель возрастает. Кислотное число выражается в количестве КОН (мг), необходимом для нейтрализации свободных жирных кислот, содержащихся в 1 г жира. В комбикормах для лососевых рыб, наиболее чувствительных к окисленным жирам, стандартный показатель перекисного числа — от 30 до 55 мг КОН/г жира, причем первый, более жесткий — для молоди [Картавцева и др., 1987].
Существует ряд способов предупреждения развития перекисного окисления липидов в комбикормах и устранения неблагоприятного воздействия продуктов окисления на организм рыб. К ним относится введение в процессе изготовления комбикормов жировых добавок с антиоксидантами естественного происхождения (например, α-токоферола, витамина С), или синтетического (ионола, сантохина, дилудина и др.), а также комплекса витамина Е с микроэлементом селеном или введение в определенных соотношениях солей цинка и меди.
Вредное влияние окисленных жиров на рыб в какой-то мере помогает снять опрыскивание комбикормов водным раствором витамина С непосредственно перед скармливанием [Князева, 1979].