Скорость синтеза белков различных органов у рыб так же, как и у высших позвоночных, неодинакова, что связано со специфическими функциями систем органов. Белки полостных органов вследствие их высокой метаболической активности синтезируются и обновляются значительно быстрее, чем белки мышц. Это подтверждается данными табл. 1 [Mambrini, Guillaume, 1999], согласно которым скорость синтеза белков в различных тканях и органах рыб существенно различается.

В количественном отношении основная масса белка откладывается в белых мышцах. По литературным данным [Hulihan et al., 1986] в мышцах радужной форели откладывается и используется на прирост массы 76% синтезированного этой тканью белка, а в жабрах — лишь 4%. У трески (при максимальном росте 2%/сут.) на прирост используется 60% белка, синтезированного в мышцах, и только 10-30% белка жабр, кишечника и желудка.
Между скоростями синтеза и распада белков обнаружена прямая корреляция. При голодании рыб наиболее быстро расщепляются и утилизируются в эндогенном питании белки белой мускулатуры. Красная мускулатура используется более медленно, что связано с ее важной функциональной ролью в процессе плавания. Белки жизненно-важных органов, в частности нервной ткани и сердца, организм оберегает [Щербина, 1989]. Интересно отметить, что у рыб при равной скорости роста, отложение протеина в белых мышцах идет быстрее, чем у крыс, и он в больших количествах используется на рост (50-70 против 25-40%).
Белки в отличие от жиров, углеводов и витаминов не откладываются в запас, они являются структурным элементом тканей. Их расход в организме при недостаточном поступлении с пищей или голодании приводит к разрушению протоплазмы клеток и, в первую очередь, клеток мышц и печени. Поэтому количество и качество белка, степень его переваримости и усвоения в организме животного во многом определяют питательную ценность корма.
Организм использует для питания не сам белок, а его структурные элементы — аминокислоты, поэтому полноценность пищевых белков во многом зависит от их химического состава, то есть набора и количественного соотношения аминокислот, а также их доступности для организма в процессах переваривания.
Белки животного и растительного происхождения обычно состоят из 20 основных аминокислот. Поэтому их называют протеиногенными. Десять из них (лизин, аргинин, гистидин, треонин, лейцин, изолейцин, валин, метионин, триптофан и фенилаланин) относят к незаменимым (эссенциальным) аминокислотам, так как они не могут синтезироваться в организме из других веществ и обязательно должны поступать с пищей, а если и синтезируются, то со скоростью, недостаточной для удовлетворения его потребностей. К заменимым аминокислотам относятся глютаминовая и аспаргиновая, серии, глицин, а-аланин, пролин, тирозин, цистин, цистеин и ряд других. В то же время при недостатке цистина на его синтез используется незаменимый метионин, а тирозина — фенилаланин. В результате потребность организма в метионине и фенилаланине возрастает. Поэтому иногда цистин и тирозин называют условно незаменимыми аминокислотами, а потребность в серусодержащих и ароматических аминокислотах выражают в виде суммы метионина с цистином и фенилаланина с тирозином [Аое et al., 1970; Nose et al., 1974; Halver, 1975; Wilson, 1984, 1989; Tacon, Cowey, 1985 и др.].
Если по качественному составу незаменимых аминокислот белки кормов в основном одинаковы, то по их количественному соотношению резко различаются. Специфические проявления недостаточности отдельных незаменимых аминокислот в питании рыб, как и других животных, не имеют в большинстве случаев яркой клинической картины и чаще всего характеризуются признаками, которые проявляются при неполноценном питании вообще. Эти признаки были обнаружены при проведении специальных экспериментов [Halver et al.,1964; Mertz, 1969; Nose,1978; Wilson, 1984,1989; Luquet, 1991 и др.]. К ним относятся замедление роста, потеря аппетита, снижение резистентности (общей устойчивости) организма.
Недостаток незаменимых аминокислот в рационах приводит к повышенном)' потреблению белка, что значительно увеличивает затраты корма на единицу прироста рыб. Поэтом)' корма по содержанию незаменимых аминокислот делятся на полноценные и неполноценные.
При резком недостатке незаменимых аминокислот в пище вступает в силу закон "минимума", согласно которому дефицит лишь одной из незаменимых аминокислот ограничивает не только эффективность использования других аминокислот, но и всего белка рациона в целом. При этом тормозится образование мышечной ткани, избыточные аминокислоты используются на энергетические нужды или после дезаминирования превращаются в жиры, или же выделяются из организма с конечными продуктами азотистого обмена (в основном через жабры в виде аммиачных солей).
Потребность в белках корма, по сути, является потребностью в аминокислотах, их качественном наборе и количестве. Естественно, что потребности в отдельных незаменимых аминокислотах у разных видов рыб могут быть неодинаковыми, поэтому в пище должно существовать такое их соотношение, которое наилучшим образом отвечает пластическим и функциональным нуждам организма. Это соотношение обычно называют "идеальной аминограммой" или "идеальным белком".
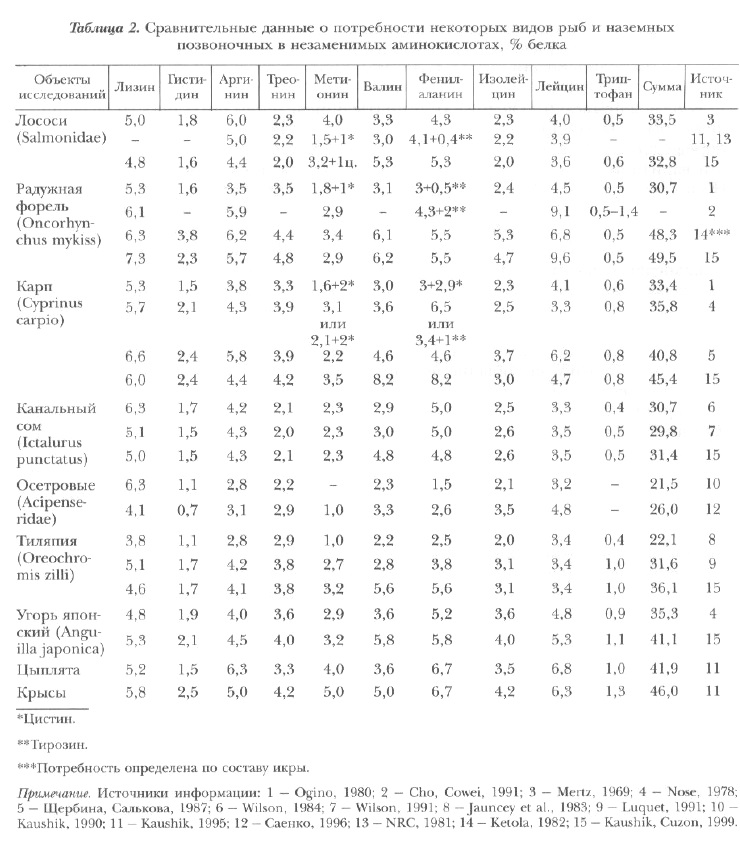
В табл. 2 приведены сводные результаты определений потребности различных видов рыб в незаменимых аминокислотах. Их анализ свидетельствует о наличии определенных вариаций. Однако в настоящее время трудно судить, являются ли эти вариации следствием видоспецифических или методологических различий. Но все же привлекает внимание их однородность для большинства аминокислот и сходство с потребностями в аминокислотах птиц и млекопитающих (крыс).
В полноценных для рыб белках корма содержание незаменимых аминокислот обычно составляет 35-50%, остальная часть приходится на долю заменимых. В жизнедеятельности и питании рыб они также имеют большое значение, так как являются поставщиками основного количества обменного азота. При их недостатке идет повышенный распад незаменимых аминокислот, в связи с чем снижается продуктивное действие пищи в целом. Однако их соотношения между собой не имеют такого значения, как соотношения между незаменимыми аминокислотами, поэтому они обычно не нормируются. В процессах обмена в ходе различных реакций заменимые аминокислоты способны к взаимопревращениям, например, путем переаминирования (обратимого переноса) аминогруппы без промежуточного образования аммиака. Другая часть аминокислот теряет аминогруппу путем дезаминирования, в результате которого образуются углеродные скелеты (кето- и оксикислоты). Последние могут превращаться в углеводы или жиры, или частично использоваться для синтеза других аминокислот путем присоединения к ним свободной аминогруппы. При другой реакции распада белка — декарбоксилировании — от его молекулы отщепляется карбоксильная группа, которая также может быть использована для синтеза углеводов и жиров.
Оценку питательности кормовых белков принято давать по тем незаменимым аминокислотам, которые содержатся в наименьших количествах и носят название лимитирующих. Для определения степени аминокислотной полноценности белка используют понятие "аминокислотный скор". Он представляет собой отношение процентного содержания незаменимой аминокислоты в корме к проценту этой же аминокислоты в "идеальном белке":
Sак= (%Aк-ты в кормовом белке/%Aк-ты в идеальном белке)·100%
За идеальное принимается то соотношение и количество аминокислот, которое удовлетворяет потребности организма и обеспечивает его оптимальный рост при минимальном уровне потребляемого белка.
В практике кормления обычно используют термин "сырой протеин". Понятие сырой протеин объединяет группу азотсодержащих веществ белковой и небелковой природы (амины, амиды, аминокислоты, пуриновые и пиримидиновые основания, аммонийные соединения и т.д.). Основу сырого протеина составляют белки. Обычно его рассчитывают путем умножения содержания общего азота в корме на коэффициент 6,25.
Потребности в общем количестве сырого протеина имеют большие видоспецифические и возрастные различия. Они изменяются в зависимости от стадии развития рыб, экологических условий (в частности, температуры, кислородного и гидрохимического режимов и состояния здоровья рыб). Быстрорастущая молодь — личинки и мальки, нуждаются в большем количестве белка, чем взрослые рыбы. Для половозрелых особей уровень белка в корме должен быть ниже, чем для ювенильной молоди, что связано с относительным увеличением расходов на поддерживающий обмен и сокращением трат на процессы роста.
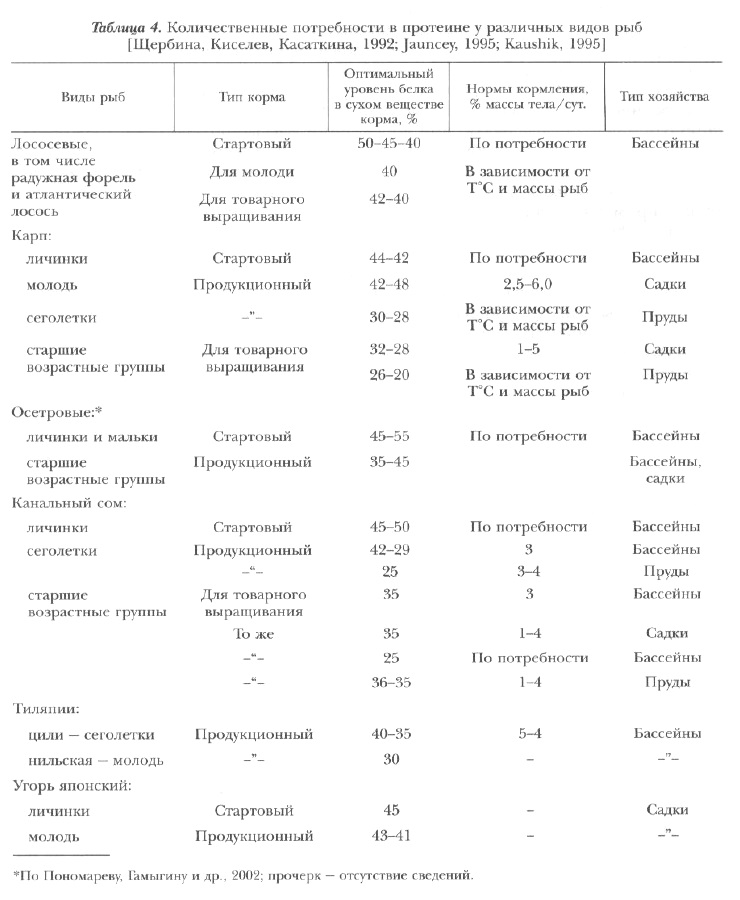
По усредненным данным, оптимальный уровень протеина в сухом веществе комбикормов для молоди всех видов рыб составляет 45-55%. Это связано со сходством питания большинства видов в естественных условиях — преимущественно животными организмами. Именно эта пища способна обеспечить потребности быстрорастущей молоди в структурных элементах для построения тканей и органов. Причем для ранних стадий постэмбрионального развития рыб белок корма частично должен быть представлен мелкими фракциями (свободными аминокислотами, ди-, олиго- и полипептидами, низкомолекулярными растворимыми белками) и в соотношениях, близких к их соотношениям в зоопланктоне.
По мере роста рыб все более проявляется видовая специфичность. Для хищных лососевых и осетровых содержание протеина в полноценных продукционных комбикормах должно составлять 34-45%, для канального сомика — 30-40%, для полифага карпа — 23-40%.
Существует зависимость между массой рыб и их требованиями к уровню протеина в корме, соблюдение которых обеспечивает максимальный рост рыб в оптимальных условиях среды. По данным И.Н. Остроумовой [1983, 1983а] при выращивании карпов в индустриальных тепловодных хозяйствах при температуре 25-30°С соотношение между массой рыб и уровнем белка в корме должно отвечать указанному ниже:

В то же время при выращивании карпов в прудах рацион является смесью комбикорма и компонентов естественной кормовой базы (зоопланктона, бентоса, детрита и т.д.), и молодь карпа массой свыше 1 г способна даже при более низких температурах хорошо расти, питаясь комбикормами с 28-30% белка (в сухом веществе). С ростом рыб их возможности удовлетворять свои потребности в белке расширяются. Карп приобретает способность питаться кормами с различными уровнем и качеством белка, набирая при этом необходимое его количество за счет увеличения потребления кормов. Молодь массой 10-15 г хорошо растет на комбикормах с 26-30% белка, при достижении массы 100-200 г — с 23-26%, более 200 г — с 20-23%. В комбикормах для двухлеток и трехлеток содержание белка может быть снижено до 18-19% при хорошей обеспеченности естественной пищей. При ее обилии и плотности посадки двухлеток в пруды менее 3000 экз/га можно применять зерна злаков, содержащие 12-13% белка [Щербина, Сапаров, Раденко, 1992].
К факторам среды, существенно влияющим на потребность рыб в белке, относится температура. У рыб как у пойкилотермных животных она определяет интенсивность синтетических процессов и скорость роста. В связи с этим требования различных видов к уровню белка в корме неодинаковы и зависят от степени удаленности температуры окружающей среды от эволюционно выработанного температурного оптимума. По сведениям Ф. Дикушниковой с соавторами [1976], оптимальная температура, при которой была установлена максимальная скорость синтеза белка у ювенильных карпов (при выращивании в садках на теплых водах), составляла около 29°С. И. Н. Остроумова [2001], обобщив собственные и литературные данные, пришла к выводу, что оптимальный рост карпов в садках при различных температурах может быть обеспечен при уровне протеина в корме, указанном в табл. 3.
При культивировании различных видов лососевых на многочисленных примерах показано, что оптимальный уровень сырого протеина в стартовых кормах при 15°С находится в пределах 52-55% (сухого вещества), при 8°С — 40-42% [De Long et al., 1958; Halver, 1970; Гамыгин, 1987].
Содержание кислорода в воде также относится к факторам, влияющим на потребность рыб в белке, и как следствие, на его относительное содержание в комбикормах. Концентрация кислорода ниже оптимальных границ ухудшает условия метаболизма и приводит к снижению эффективности использования белка рыбами. В результате ростовой и продукционный эффекты при оптимальном (для данной температуры и массы рыб) и меньшем на ¼-⅓, относительном содержании белков в корме могут сближаться. Аналогичное действие могут оказывать загрязнение воды и состояние здоровья рыб.
Оптимальный уровень сырого протеина в кормах зависит от качества используемых белков (чем хуже качество, тем выше должно быть содержание белка) и предполагаемых норм кормления. Последние определяются способностью рыб поедать и переваривать определенный объем корма. В качестве примера в табл. 1 Приложения приведены потребности карпа в белке при различных температурах культивирования и нормы кормления рыб в зависимости от их массы и относительного содержания белка в комбикорме при выращивании в индустриальных тепловодных хозяйствах.
Показано, что оптимальный уровень белка в комбикормах находится в определенной связи с энергетической обеспеченностью рациона. Если ранее полагали, что наибольшей эффективностью для лососевых обладают корма, где доля энергии белка составляет 40-65%, то в дальнейшем было установлено, что белок может быть сэкономлен, если корм будет обеспечен достаточным количеством энергии за счет безбелковых источников. Наибольшую ценность в этом отношении представляет жир. Высокая эффективность выращивания радужной форели на протяжении всего жизненного цикла была получена при использовании корма с относительно низким содержанием протеина и высоким — жира, соответственно 35-38 и 30-36% [Cho, Kaushik, 1985, 1990].
Меньшую энергетическую ценность представляют углеводы, в частности сырой крахмал, обладающий низкой переваримостью.
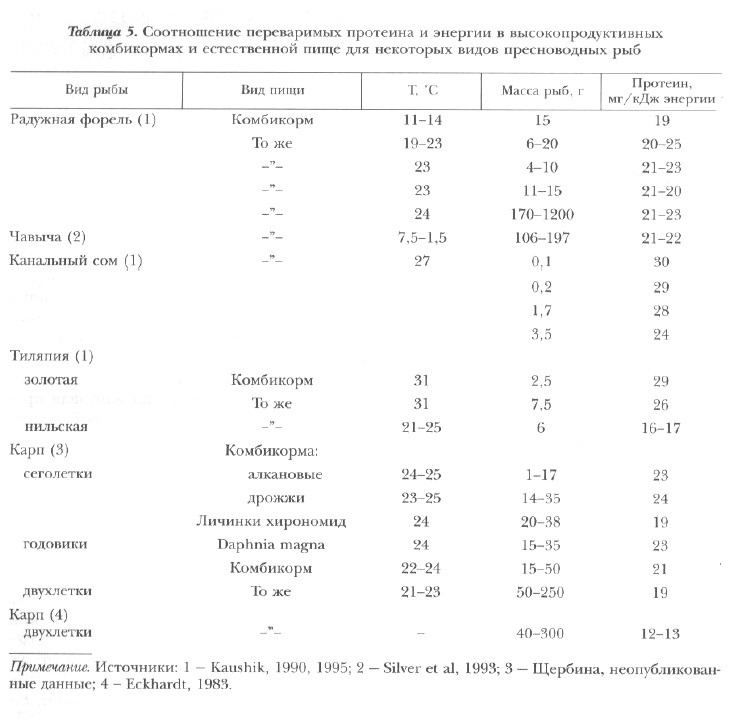
Оптимальные соотношения протеина и энергии для большинства объектов аквакультуры находятся в пределах 12-30 мг переваримого протеина на 1 кДж переваримой энергии, а протеина и жира — 36:18 [Kaushik, 1990] (табл. 5).