
По количественному содержанию в животных и растительных тканях они делятся на макро- и микроэлементы. К макроэлементам относят кальций, фосфор, магний, калий, натрий, хлор, серу. Их количество в организме рыб колеблется в широких пределах, составляя более 100 мг/кг. К микроэлементам относят железо, медь, марганец, цинк, кобальт, селен и другие (содержание в пределах 0,01-90 мг/кг).
В среднем общая сумма минеральных элементов в теле рыб составляет 2,5-8,0%, причем на долю макроэлементов приходится более 99,5%.
Особенностью минерального питания рыб является наличие двух путей поступления минералов в организм рыб: с пищей через рот и посредством осмотического проникновения из воды через жабры и покровные ткани.
Доказано [Карзинкин, 1962; Шеханова, 1956], что рыбы способны активно сорбировать из воды кальций, магний, натрий, калий, фосфор, серу, хлор и другие элементы. Поэтому потребность рыб в минеральных веществах, содержащихся в воде в высоких концентрациях, может удовлетворяться в значительной мере или полностью (например, в кальции и магнии) осмотическим путем. В зависимости от того, в какой воде ведется выращивание рыб (пресной или солоноватой, мягкой или жесткой), они вынуждены постоянно либо восполнять недостаток определенных минеральных веществ, либо экскрегировать их избыток. Качественный и количественный состав минеральных солей в различных водоемах неодинаков, что обусловливает разную степень их абсорбции из воды и различия в требованиях организма рыб к минеральному составу кормов. Следовательно, солевой состав воды может существенно влиять на минеральный обмен у рыб. Элементы, концентрация которых в воде обычно невелика, должны поступать в организм с пищей. Обычно к ним относят фосфор, цинк, йод, марганец, кобальт, селен и др.
Помимо воды, источником минералов для рыб служат комбикорм и естественная пища.
В связи с особенностями компонентного состава комбикормов для прудовых карповых рыб (злаковые, жмыхи и шроты масличных культур, кормовые дрожжи и незначительные количества рыбной или мясокостной муки), они бедны минеральными элементами. Кроме того, минеральные вещества ряда сырьевых компонентов могут находиться в форме, которая плохо доступна пищеварительным ферментам безжелудочных рыб (например, фосфор рыбной муки в виде гидроксиапатита или фосфор злаковых в соединении с фитиновой кислотой, а также фитаты железа, цинка, марганца). Комбикорма для форели и других видов рыб, содержащие значительно большее количество рыбной муки и другого животного сырья, более богаты минералами. Однако и у этих рыб, несмотря на присутствие в желудках соляной кислоты, значительная часть фитатов многих элементов, а также гидроксиапатиты костной ткани, перевариваются недостаточно хорошо.
Естественная пища (в частности, планктон, бентос, насекомые, подводная растительность, рыбы) содержит все необходимые элементы в физиологически согласованных соотношениях в соответствии с солевым составом воды, и поэтому она является важным дополнением, нивелирующим недостатки минеральной части комбикормов.
Ограниченное или избыточное поступление минеральных веществ в организм рыб, обусловленное особенностями состава комбикормов, может привести к снижению аппетита, возникновению патологических изменений (особенно на ранних стадиях развития) и торможению роста. Например, при недостаточности в рационе фосфора, магния, марганца, цинка, кобальта в совокупности или хотя бы одного из этих элементов развивается остеодистрофия, которая проявляется в различных искривлениях позвоночника, деформации лобных и челюстных костей, ротового аппарата, редукции жаберных крышек, искривлениях ребер и в ряде других патологий, в частности, в жировом обмене. Все эти изменения тормозят рост рыб и снижают их продуктивность.
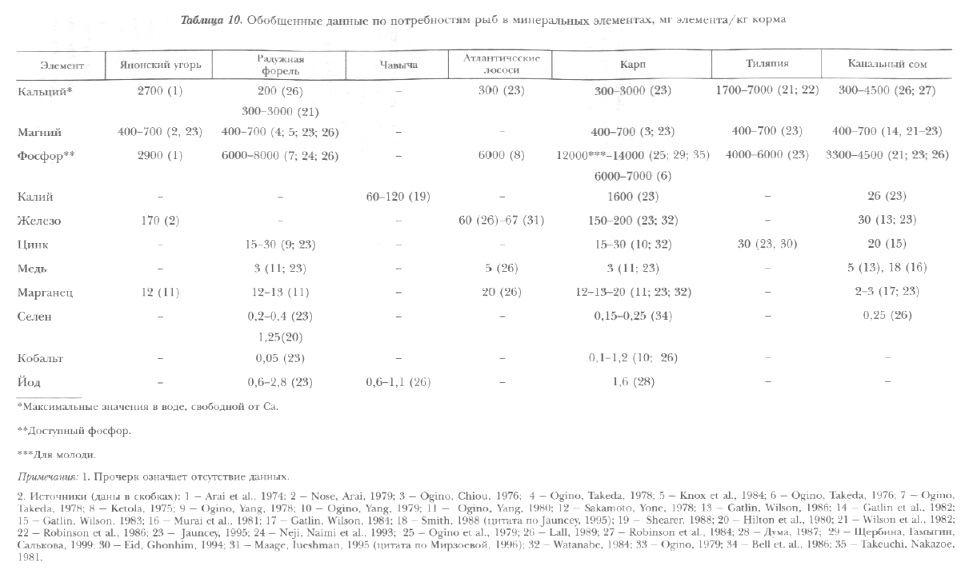
Количественные потребности рыб в минеральных элементах в связи с особенностями их поступления в организм изучены недостаточно полно (табл. 10). Как правило, их определяли на молоди в сериях специальных экспериментов при относительно постоянном составе воды.
Отдельные соли и их смеси вводили в экспериментальный корм одного состава, достаточно полноценный по всем остальным питательным веществам. Потребность оценивали как минимальное количество элемента, обеспечивающее быстрый рост молоди рыб и нормальное содержание элемента в скелете, печени и мышцах.
Однако чувствительность рыб к содержанию макро- и микроэлементов в комбикормах в реальных ситуациях иная. Она определяется комплексом факторов: концентрацией и соотношением солей в воде, качественным и количественным соотношением компонентов в комбикорме, формой солей, в которых они находятся в кормах, доступностью их для организма. Немаловажное значение имеет также степень обеспеченности рациона другими питательными веществами (белками, жирами, углеводами), необходимыми для нормального обмена веществ. Кроме того, потребность в минеральных элементах зависит от температуры. При ее повышении активизируются обменные процессы и возникают условия для ускорения роста рыб, поэтому' требования организма к минеральному' составу корма возрастают, а в случае их недостаточного удовлетворения быстрее проявляются патологические изменения. Признаки недостаточности отдельных минеральных веществ приведены в табл. 11.
Ознакомимся более подробно со значением и ролью в организме рыб основных минералов.
Кальций по количественному содержанию в теле рыб занимает первое место. Он концентрируется в костях скелета (около 80%), чешуе (~ 12%) в мелких костях (~ 4-5%) и отолитах (3-4%); в мягких тканях его содержится около 1-2% [Ichii, Mugiya, 1983].
В организм рыб кальций может поступать как из воды, так и из пищи, при этом доминирует первый, осмотический, путь. Причем 90% осмотического кальция проникает через жабры, остальное — через кожу, плавники и ротовой эпителий.
Кальций является главным элементом минерального вещества кости и обеспечивает ее прочность. Он участвует в процессах регуляции проницаемости клеточных мембран, проведении нервного импульса, сокращения мышц и свертывания крови. Играет активную роль во множестве других биохимических реакций, происходящих в организме. Потребность в кальции зависит от вида, возраста и массы рыб. Молодь нуждается в большем количестве кальция, чем рыбы старших возрастных групп. Максимальная потребность в нем отмечена в период окостенения скелета и развития чешуи (например, у молоди карпа массой около 100 мг). Экспериментально определенная потребность в пищевом кальции колеблется в широких пределах, что зависит от его концентрации в воде. Для карпа и радужной форели это составляет 0,3-3,0 г/кг корма, гиляпии — 1,7-7,0 г/кг (максимум — при ‘минимуме в воде). В опытах с радиоактивными изотопами установлено, что при содержании кальция в воде, равном 30 мг/л потребность в нем полностью может удовлетворяться осмотическим путем [Карзинкин, 1962]. При этом чем больше кальция растворено в воде, тем выше возможность его поступлении путем осмоса. Поэтому необходимость в пищевом кальции для удовлетворения потребности возникает при низкой концентрации его в воде, что имеет место в хозяйствах со слабо минерализованным источником водоснабжения. В то же время следует иметь в виду, что избыток кальция в корме, как и его недостаток, способен привести к нарушению минерального обмена и, как следствие, к замедлению роста и снижению оплаты корма. Например, избыточное поступление кальция с кормом в организм карпа при одновременно высоком содержании его в воде приводит в экскреции этого элемента из организма через пищеварительную систему [Щербина, 1973; Казлаускене, Щербина, 1975]. Аналогичное явление наблюдалось и у канального сома при 2% Са в корме [Gatlin, Phillips, 1989].
Одной из распространенных минеральных добавок к комбикормам для прудового карпа является мел (СаСО3, который вводится в количестве 1-2%. Однако эффективность этих добавок определяется содержанием кальция в воде. Его положительное влияние наблюдается только при концентрации до 40 мг Са++/л и жесткости воды, не превышающей 10 нем. град. При концентрации более 50 мг/л для сеголеток и 40 мг/л для двухлеток и трехлеток карпа присутствие мела в комбикормах вследствие нарушения деятельности пищеварительной системы и обмена веществ приводит к замедлению роста, уменьшению накопления питательных веществ в теле рыб и повышению кормовых коэффициентов. При более высоких концентрациях кальция в воде добавки мела вызывают резкое торможение роста рыб [Казлаускене, Щербина, 1973а]. Сходное действие на карпа оказывал уровень кальция 0,5-2,0%, создаваемый смесью солей СаСО3 и СаС12·Н2O. У канального сома при концентрации кальция в воде 100 мг/л отмечалось [Gatlin, Scarpa, 1993] торможение роста и снижение эффективности кормления. В то же время исследования на форели, выполнявшиеся при рециркуляционном водообеспечении, показали, что независимо от концентрации кальция в воде его содержание в корме 2-3 г/ кг оказывает ростостимулирующее действие [Сергеева, 1989].
Большое влияние на усвоение кальция и рост рыб оказывают обеспеченность корма витамином Д, а также соотношение поступающих в организм кальция и фосфора. У различных видов рыб это соотношение достаточно вариабельно (от 1:1 до 1:2), в частности, для карпа оно составляет 1:1 [Hilton, 1978, цит. по Kaushik, 1990]. Для большинства других видов оно приближается 1:2 (например, содержание кальция в корме — 0,34%, фосфора — 0,68%). При этом фосфор должен находиться в доступной для организма рыб форме, что будет обсуждаться ниже. При недостатке кальция в корме и достаточном содержании в нем фосфора возникающий дефицит кальция может полностью компенсироваться за счет его осмотического поступления из воды.
Фосфор в теле культивируемых рыб обычно содержится в количестве от 0,7 до 1,2%. Причем около 80% находится в скелете, так как вместе с кальцием он входит в минеральный остов кости (кристаллы гидроксиапатита — Са10(РО4)6(ОН)2). Остальное количество фосфора распределяется в мягких тканях и входит в состав разнообразных органических соединений (нуклеопротеидов, ферментов, ко-ферментов, АТФ, АДФ, фосфолипидов и т.д.). Вместе с ними он участвует в процессах переваривания и всасывания питательных веществ, биологического окисления, гормональной регуляции, обеспечивает буферные свойства крови. Являясь одним из составных элементов нуклеиновых кислот, он принимает участие в передаче наследственной информации. Фосфор фосфолипидов служит важным компонентом всех клеточных мембран, отделяющих внутреннее содержимое клеток и их органелл от окружающей среды.
Признаки недостаточности фосфора у рыб проявляются в разной мере. У рыб старших возрастных групп наблюдается снижение аппетита, уменьшение переваримости и усвоения корма, в частности белка, угнетение роста, торможение синтеза белка при одновременном ожирении [Ogino, Takeda, 1976; Ogino et al., 1979]. У двухлеток карпа при сверхвысокой плотности посадки в прудах (10-30 тыс. экз/га) и практическом отсутствии естественной пищи нередко отмечаются искривление позвоночника и снижение продуктивного действия корма. Иногда наблюдается недоразвитие костей и бледность внутренних органов [Nose, Arai, 1979]. У личинок и молоди недостаток фосфора тормозит развитие, приводит к искривлениям позвоночника (лордозу, кифозу), возникновению деформаций черепа, среди которых наиболее заметно изменение лобной кости. Наблюдаются нарушение кальцификации ребер, накопление в теле жира, снижение содержания воды и минеральных элементов. Сеголетки карпов, выросшие на комбикормах, дефицитных по фосфору, в условиях высокой плотности посадки (45-60 тыс. экз/га) и слабой обеспеченности естественной пищей при зимнем голодании менее жизнеспособны и несут большие энергетические потери, даже если достигли стандартной массы тела и высокой жирности. Особенно большой урон недостаток фосфора в комбикормах наносит в индустриальных хозяйствах с рециркуляционной системой водоснабжения, при которой отмечается повышенное содержание кальция в результате избирательного питания микрофлоры биофильтров.
Потребности в фосфоре, так же как и в других элементах, зависят от возраста, физиологического состояния рыб, температуры среды и состава комбикорма. Они максимальны у личинок и молоди в период окостенения скелета (при массе 0,1-1,0 г). В случае питания рыб кормами с высоким содержанием белка (45-35%) потребность в фосфоре выше, чем с низким — 26-20%. Доступность фосфора, содержащегося в различных видах сырья, и минеральных солей для организма желудочных и безжелудочных рыб неодинакова (см. раздел 2.4.4.). Поэтому в последнее время потребность организма рыб в этом элементе выражается с учетом показателя его доступности, характеризующего способность пищеварительной системы извлекать фосфор из различных кормов и минеральных добавок к ним. Например, для быстрорастущих личинок и молоди карпа массой до 12 г, количество доступного фосфора в корме должно быть не менее 12-14 г/кг, для рыб растущих медленнее, а также для холодноводной форели — около 7-8 г/кг, лосося — 6 г/кг; для карпа старших возрастов, форели, тиляпии, канального сомика достаточно 6-4 г/кг [Ogino, Takeda, 1976; Takeuchi, Nakazoe,1981; Nakamura,1982; Tacon, De Silva, 1983; Щербина, Чяпулис, Гамыгин, 2000].
По данным И.А. Шехановой [1956], для всех видов рыб фосфор естественной пищи доступен хорошо — до 85%. В то же время фосфор рыбной муки, представленный в основном гидроксиапатитом костной ткани, личинками практически не усваивается. Его доступность для рыб старших возрастов также невелика — 10-25%.
Доступность фосфора стандартных комбикормов рецептов К-110, К-111 и К-112 для сеголеток и двухлеток прудового карпа составляет около 30-40%. В силу того, что эти комбикорма состоят в основном из растительного сырья, бедного фосфором, его дефицит может быть покрыт либо за счет обеспечения рыб естественной пищей, либо за счет дополнительного введения в комбикорма легкодоступных фосфорных солей. С этой целью применяют различные минеральные фосфаты. Для карпов старших возрастов хороший эффект дает применение 1-3% диаммоний- и дикальцийфосфата. Содержание общего фосфора в дикальцийфосфате составляет 228 r/кг, доступного — 105 г/кг. Введение его в комбикорм дает повышение прироста рыб на 20-40% при сокращении затрат кормов. Добавки трикальцийфосфата (около 200 г/кг общего и 26 г/кг доступного фосфора) не желательны, так как чаще всего они дают отрицательный эффект, создавая в корме нежелательный избыток кальция. Они могут оказывать благоприятное влияние только при низком содержании кальция в воде (менее 30 мг/л) [Щербина, Гамыгин, Чяпулис, 2000]. В случае выращивания личинок с использованием стартовых комбикормов "Эквизо" и РКС хорошее действие оказывают такие добавки, как однозамещенные фосфаты калия, натрия и кальция или их смеси, являющиеся хорошо доступными для рыб источниками фосфора (усвояемость более 90%). Их вводят в количестве 1-3%. Это ускоряет рост личинок (на 20-50%), существенно повышает их выживаемость (в 1,3-2,0 раза) и эффективность использования комбикормов. Причем этот эффект проявляется тем сильнее, чем выше температура, при которой идет выращивание личинок, и чем интенсивнее рост [Аршавский, 1987].
В НТЦ "Аквакорм" был апробирован специальный премикс к стартовым комбикормам для личинок карпа, предупреждающий развитие скелетных аномалий. Его состав на 100 г корма: КН2РO4 — 3 г (680 мг Р), Zn·SO4·7H2O — 88 мг (20 мг Zn), MnCl2·4Н2O — 36 мг (10 мг Mn), витамин С — 500 мг (или в виде полифосфата — 150 мг). Рыбоводный эффект применения комбикорма РКС-ЗМ, обогащенного этим премиксом, при выращивании личинок карпа в проточных и рециркуляционных установках выражался в следующем: по сравнению с контролем были отмечены снижение скелетных уродств с 70-60 до 20-14%, увеличение выживаемости личинок в 2,0-2,6 раза; ускорение темпа роста рыб в 1,5-2,5 раза; повышение выхода продукции, снижение затрат корма на прирост в 1,5-2,0 раза [Щербина, Гамыгин, Салькова, 1992].
Добавки в комбикорма моно- и дифосфагов калия, натрия, аммония в количестве 1-3% оказывают благотворное влияние и на молодь карпов (массой 1-10 г и более). В специальных 30-суточных экспериментах НТЦ "Аквакорм" было показано, что оно выражается в сокращении общей потребности в пище на 16-32% и одновременном увеличении потребления рыбами доступного фосфора с 16 до 74 мг на 100 г массы рыб. При этом обнаружено увеличение общей переваримости корма на 3-15% в основном за счет повышения абсорбции белка (до 13%) и минералов (на 150-200%). Кормовые коэффициенты снижались на 19-39%. За счет активизации метаболизма происходило ускорение роста рыб на 20-50%. Максимальный эффект оказала добавка в корм монокалий фосфата (КН2РO4) в дозе 3% [Щербина, Гамыгин, Чапулис, 2000].
Исходя из этого, при наличии ассортимента рецептур стартовых комбикормов для личинок и возможности выбора следует отдавать предпочтение тем рецептам, которые содержат фосфорные добавки, несмотря на их более высокую стоимость. В равной степени это относится и к комбикормам для сеголеток и старших возрастных групп карпа, особенно если планируется выращивание в индустриальных условиях или в прудах при уплотненной посадке.
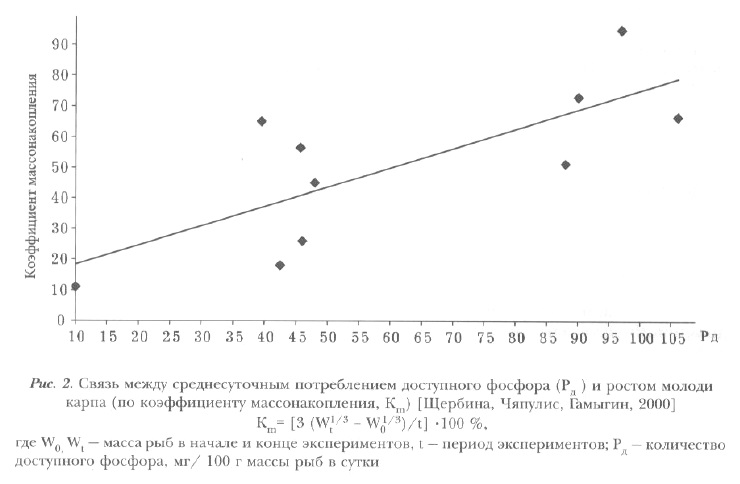
РИСУНОК На рис. 2 представлена зависимость между ростом молоди карпа, выраженным коэффициентом массонакопления, и среднесуточным потреблением доступного фосфора.
Для рыб, имеющих желудок и солянокислое пищеварение, в частности форели и др. лососевых, фосфор рыбной муки доступен значительно лучше. Однако фосфор фитатов усваивается ими также плохо, как и безжелудочными. Поэтому этот тип комбикормов, как правило, также следует обогащать фосфатами.
Магний входит в состав тела рыб приблизительно в количестве 0,3%. Более половины магния сосредоточено в костной ткани, где он в совокупности с кальцием и фосфором (в виде фосфорнокислого магния) образует минеральный остов скелета и его присутствие повышает прочность костной ткани. Около 40% магния распределено в мягких тканях. Здесь он активизирует некоторые ферменты окислительного фосфорилирования и цикла Кребса и является составной частью ряда ферментов. Кроме того, магний играет важную роль в обмене нуклеиновых кислот, обеспечивает функционирование нервно-мышечного аппарата, участвует в осморегуляции.
Значительная часть магния поступает в организм рыб осмотическим путем, однако при концентрации его в пресной воде менее 5 мг/л рыбы должны получать его с кормом. Количество магния, необходимого для нормального развития и быстрого роста различных видов рыб, определено в пределах 400-700 мг/кг корма. Для молоди карпа массой до 100 мг максимальная потребность в магнии составляет 0,6-0,7 г/кг корма. До достижения массы 1 г в период формирования скелета она остается такой же. С возрастом потребность уменьшается: так при массе рыб от 1 до 10 г она составляет 0,3-0,4 г на 1 кг корма, при массе от 10 до 100 г и более — приблизительно 0,2 г/кг. С повышением температуры окружающей среды потребность в магнии возрастает. Избыточное количество в корме кальция и фосфора увеличивает потребность организма рыб в магнии, что необходимо для нормального формирования скелета [Ogino, Chiou, 1976].
В растительных компонентах комбикормов магний может находиться в форме фитатов. В этом случае его доступность для рыб может колебаться в широких пределах (35-87% у форели [Sugiura et al., 1998]). Магний рыбной муки, содержащей много костей, доступен слабо — 26%, т.к. он находится в форме гидроксиапатитов. При малом содержании костей в муке его доступность возрастает до 60-64%. Резкий недостаток магния в корме приводит к симптомам, возникающим при дефиците многих других пищевых веществ: снижению аппетита и торможению роста рыб, а также к различным патологиям (см. табл. 11).
У прудовых рыб благодаря особенностям компонентного состава комбикормов и присутствию в их рационах естественной пищи недостаточность магния в комбикормах, сопровождающаяся патологией, может проявляться только в хозяйствах на слабоминерализованных водах и в период повышенных температур. В индустриальных хозяйствах она вполне вероятна. Для предотвращения патологий необходим контроль за содержанием магния в воде и соотношением в ней магния и кальция. Известно, что повышенное содержание в воде кальция при низком содержании магния препятствует проникновению последнего в организм. Оптимальное соотношение кальция и магния в воде 3,7:1,0 [Карзинкин, 1962].
Цинк относится к микроэлементам. Он сосредоточен в основном в костях, печени, жабрах. Много цинка в глазах и гонадах, что связано с его важной ролью в функционировании органов зрения и развитии половых продуктов. Цинк поступает в организм рыб как из корма, так и из воды. Высокое содержание цинка в воде (до 0,3 мг/л против ПДК 0,01 мг/л), часто вызванное загрязнением водоемов промышленными отходами, может оказывать токсическое воздействие на организм рыб. Патологическое накопление цинка в теле (например, в мышцах до 20-45 мг/кг против 1-6 мг в норме) приводит к торможению процессов, которые при малых концентрациях он обычно активирует (кроветворение, зрение, костеобразование, размножение, развитие, рост).
Потребность в пищевом цинке для карпа, форели, канального сомика и тиляпии составляет 15-30 мг/кг. Недостаточность цинка в кормах выражается в потере аппетита, торможении роста, укорочении тела, снижении жирности, а также в повышении смертности (см. табл. 11). Специфическими признаками дефицита цинка в пище являются воспаление и эрозия плавников и кожи, а также снижение содержания цинка в костях. Последнее значительно раньше свидетельствует о дефиците цинка в корме, чем ослабление роста. У молоди при недостатке цинка в кормах может возникнуть А-гиповитаминоз, который не устраняется даже при избытке каротина. Наиболее остро эта недостаточность проявляется в индустриальных хозяйствах при подращивании личинок на стартовых комбикормах при температурах, обусловливающих высокий темп роста рыб. Это связано с очень слабой доступностью цинка, входящего в состав различных видов кормовой рыбной муки (в основном белой) с высоким содержанием золы (17-22%), где он находится в виде малорастворимых солей.
Кроме того, большинство других компонентов комбикормов, в частности злаковые и масличные культуры, имеют в своем составе фитиновую кислоту. Помимо фосфора, она связывает и цинк, делая его недоступным для лососевых, карповых, сомовых и других видов рыб, так же как и для организма моногастричных животных. К примеру, для канального сома при наличии в корме 0,5% фитатов требуется повышенное количество цинка — до 200 мг/кг корма (вместо 18-20 мг согласно табл. 10). Отрицательное влияние на доступность цинка, содержащегося в кормах для рыб, оказывают добавки в корм трикальцийфосфата.
У лососевых рыб дефициту цинка в кормах сопутствует развитие непаразитарной катаракты, которая представляет серьезную проблему для индустриальных хозяйств. Она характеризуется начальным помутнением хрусталика (с дальнейшим образованием язв на роговице) и глазного яблока. Опасность возникновения катаракты возрастает, когда рыбы получают корм с высоким содержанием белой рыбной муки или при введении минерального иремикса из солей кальция, фосфора, натрия и калия. Для предотвращения катаракты у радужной форели в корм рекомендуется вводить соли цинка в количестве 15 мг на 1 кг корма. Для атлантического лосося рекомендуемые дозы выше — 12-42 мг/кг продукционного корма и 67 мг/кг стартового. У личинок карпа, помимо замедления роста и гибели, основные признаки дефицита цинка — появление белой каймы на плавниках и побледнение кожи. В этих случаях корма рекомендуется обогащать сернокислым цинком из расчета 20 мг на 1 кг корма. В связи с различиями в количестве рыбной муки, вводимой в корма для форели рекомендуются [Satoh et al., 1987] добавки ZnSO4 или Zn(NO3)2, из расчета 40 мг на 1 кг рыбной муки.
Марганец принимает участие в окислительно-восстановительных процессах, тканевом дыхании, кальциево-фосфорном обмене, оказывая тем самым влияние на костеобразование и кроветворение, нормализует липидный обмен, препятствуя жировой дегенерации печени, влияет на воспроизводительную функцию. Он активизирует синтез белков, благодаря чему интенсифицируется рост рыб. Стимулирует синтез и депонирование аскорбиновой кислоты в тканях. Марганец оказывает благоприятное влияние на усвоение и расход витаминов А, Е, К и С, снижает потребность в витамине Е.
Марганец поступает в организм рыб в основном с пищей; его количество в корме должно составлять от 2-3 (канальный сомик) до 12-13 мг/кг (карп, форель, угорь). Степень доступности для организма рыб марганца, входящего в состав кормов различного происхождения, невелика. По наблюдениям Н.Т. Сергеевой [1987], радужная форель извлекала из корма РГМ-5В всего 12% марганца. Избыток в кормах кальция и фосфора, особенно в виде трикальцийфосфата или гидроксиапатига белой рыбной муки, угнетает всасывание марганца в пищеварительном тракте лососей.
Недостаточность марганца проявляется в неправильном развитии скелета и укорочении тела, или "карликовой короткотелости" карпа и форели, возникновении уродств, нарушении структуры и формы плавников (у форели — хвостового), жировом перерождении печени. Все это сопровождается снижением скорости роста рыб (карп, тиляпия, форель), увеличением смертности, снижением аппетита и эффективности кормления, низким выклевом лососевых из икры [Ogino, 1979; Satoh et al., 1983].
Дефицит марганца в корме проявляется главным образом в случае выращивания молоди рыб при отсутствии естественной пищи. Наиболее часто он возникает при заводском подращивании личинок и молоди на стартовых комбикормах, которые имеют в своем составе много рыбной муки, особенно белой. Входящий в ее состав марганец плохо доступен для рыб. В этих случаях рекомендуется его дополнительное введение в форме серно- или солянокислых солей в количестве 10-13 мг на 1 кг комбикорма.
Предельно допустимая концентрация (ПДК) марганца для воды рыбохозяйственных водоемов составляет 0,01 мг/л.
Железо рыбы получают в основном с пищей, оно играет важную роль в процессах дыхания и биологического окисления, принимая участие в переносе электронов. Как правило, железо находится в организме в виде сложных форм, соединенных с белками (гемоглобин крови, миоглобин мышц, ферменты биологического окисления — цитохромоксидазы, трансферазы, каталазы, пероксидазы и т.д.). Недостаток железа в организме вызывает железодефицитное малокровие (микроцитарную гипохромную анемию). Она характеризуется пониженным содержанием железа в крови, снижением концентрации гемоглобина, числа эритроцитов, гематокрита (общего количества форменных элементов), повышением числа молодых эритроцитов. Результат — угнетение роста рыб, снижение эффективности использования комбикорма (см. табл. 11). Подобные явления наблюдались у форели, канальных сомов, желтохвоста, угря [Lall, 1979]. Развитие заболеваний может быть предотвращено добавлением в 1 кг корма 15-20 мг железа [Robinson, 1989]. В последнее время для коррекции алиментарных анемий у рыб с успехом используется препарат ультрадисперсного железа в дозах от 0,1 до 0,5 мг/кг корма [Головина и др., 1998].
Избыток железа, создаваемый в корме присутствием больших количеств кровяной и перьевой муки, а также селезенки, может оказаться таким же нежелательным, как и недостаток. Оно катализирует образование гидроперекисей и пероксидов, способствуя перекисному окислению липидов. Введение железа в корм в виде сернокислой соли может способствовать активизации этих процессов, особенно при наличии больших количеств полиненасыщенных жирных кислот рыбьего жира и вызвать разрушение витамина С [Watanabe et al., 1997]. У атлантического лосося обнаружена связь между избытком железа в печени и гибелью рыб от фурункулеза [цит. по Мирзоевой, 1996].
Потребность рыб в железе колеблется в широком диапазоне — от 30 мг/кг корма у канального сома до 200—300 мг/кг у карпа и форели (см. табл. 10). Доступность железа для организма рыб зависит от формы его солей. ЕПо литературным данным, что двухвалентное железо абсорбируется значительно полнее трехвалентиого [Lall, 1979]. Железо комбикормов слабо доступно для организма рыб, так как значительная часть его, также как и фосфора, цинка, марганца, входит в состав фитатов, труднорасщепляемых в кишечнике. Всасывание железа у рыб может тормозить также и присутствие в корме легкорастворимых солей фосфора [Sugiura et al., 1998].
Предельно допустимая концентрация (ПДК) железа для воды рыбохозяйственных водоемов 0,005 мг/л.
Содержание кобальта в организме рыб невелико — около 0,25-0,5 мг/кг. Он является частью молекулы витамина В)2, регулирующего образование красной крови, стимулирует синтез витаминов В; и В2, предупреждая тем самым развитие анемии. Кроме того, кобальт участвует в синтезе белков, в частности гамма-глобулинов, обеспечивающих иммунитет. Он влияет на обмен азота нуклеиновых кислот и минеральных веществ, а также способствует накоплению в печени естественных антиоксидантов (витаминов Е, С) и витамина А.
Поступает кобальт в организм рыб с пищей и осмотически через жабры [Карзинкин, 1962]. Последний путь менее значим, т. к. большинство рыбоводных хозяйств расположено в биогеохимических провинциях, характеризующихся недостатком кобальта.
Дефицит кобальта в организме тормозит синтез гемоглобина, что может привести к возникновению анемии и снижению общей резистентности организма, повышению смертности при одновременном замедлении роста рыб (см. табл. 11). Поэтому еще с середины XX века делались попытки удобрения кобальтом воды прудов с целью обогащения рационов рыб этим элементом через трофические цепи, а также введения его непосредственно в комбикорма [Суховерхое и др., 1961; Шабалина, 1964, 1968].
Экспериментально определенная потребность форели в кобальте составляет около 0,05 мг/кг корма, карпа — 0,1 мг/кг (см. табл. 10). Однако увеличение его содержания до 1,2 мг/кг [Ogino, 1979] также может вызывать стимуляцию роста рыб. В отечественной практике [Суховерхов и др., 1961] было показано, что введение в корм для сеголеток прудового карпа хлористого кобальта из расчета 1-3 мг на 1 кг корма ускоряет их рост и обеспечивает повышение резистентности рыб. Последействие обогащения корма кобальтом выражалось в сокращении смертности молоди в зимний период и снижении трат организма при зимнем голодании. В то же время следует учитывать, что в этих случаях кобальт вносили в виде водных растворов в рассыпные комбикорма, которые замешивали в виде крутого теста, и поэтому он быстро экстрагировался и терялся в воде. Его фактическое количество, попадавшее в организм рыб, было во много раз меньше расчетного.
В комбикорма для лососевых его вводят в виде CoSO4 в дозах 0,2-3,2 мг/кг, а иногда и более [Маликова, 1967, Watanabe et al., 1997]. В то же время избыток кобальта вызывает патологические изменения в крови, а также приводит к замедлению роста рыб [Шабалина, 1968].
Селен также относится к микроэлементам, обладающим высокой активностью. В больших количествах он крайне токсичен. Есть сведения, что при его содержании в корме 11-15 мг/кг у атлантического лосося и молоди радужной форели наблюдали понижение аппетита, торможение роста, отложение камней в почках, гибель рыб [Lovell, 1996]. Токсические свойства селена проявлялись у форели также при длительном питании кормами, содержащими его в количестве 3 мг/кг.
В микродозах селен оказывает благоприятное влияние на ход биологического окисления в организме, стабилизирует проницаемость клеточных мембран, принимает участие в обмене ряда аминокислот. В метаболизме он тесно связан с витамином Е, тормозящим окисление ненасыщенных жирных кислот, а в составе фермента глютатион-пероксидазы защищает клетки от повреждающего воздействия уже образовавшихся перекисей.
Селен поступает в организм рыб как с пищей, так и из воды, при этом пути его метаболизма различны. Поступая из воды через жабры, селен разносится кровью по всему организму (исключая печень) и находится в тканях в основном в неорганической форме; поступив в печень, переходит в органическую форму, и в случае его избытка может выводиться из организма.
Рыбы обладают выраженной способностью аккумулировать селен из воды и корма. Однако большинство территорий России дефицитны по селену. Его избыток в почвах и в воде обнаружен только в ряде биогеохимических провинций Южного Урала и Тувы, поэтому основной источник селена для рыб — пища.
Потребность радужной форели в селене колеблется в пределах от 0,15 до 1,25 мг/кг корма, карпа — 0,15-0,25 мг (см. табл. 10). Корма, содержащие около 2% и более рыбной муки, обычно удовлетворяют потребность рыб в селене.
Основными признаками недостаточности селена являются торможение темпа роста рыб, снижение активности глутатион-пероксидазы в плазме крови и печени. Возможны также снижение гематокрита, повышение содержания в теле жира и воды, патологические изменения в клетках печени и спинного мозга. У форели наблюдался геморрагический экссудативный диатез, обусловленный нарушением проницаемости клеточных оболочек и гемолизом эритроцитов. При значительном дефиците селена возрастает смертность рыб с признаками анемии, повышенного содержания протеина в плазме и другими патологиями (см. табл. 11).
Критерием обеспеченности рыб селеном является уровень в печени и сыворотки крови глутатион-пероксидазы. Этот фермент является главным средством защиты организма от накопления в клетках Н2O2 и органических перекисей, образующихся в результате обменных реакций. Повреждающая функция Н2O2 обусловлена ее способностью атаковать и разрушать двойные связи ненасыщенных жирных кислот фосфолипидов клеточных мембран. В итоге это приводит к нарушению структуры и функций клетки и ее органелл. Наиболее часто такому повреждающему влиянию подвергаются печеночные клетки (гепатоциты), что нередко служит основной причиной часто встречающейся патологии — жирового перерождения печени. Образующиеся гидроперекиси вызывают реакции, которые приводят к разрушению оболочек эритроцитов (гемолизу) и позже к развитию гемолитической анемии. Максимальная активность глютатион-пероксидазы в плазме крови форели отмечена при содержании селена в корме в количестве 0,15-0,38 мг/кг. Для поддержания эритроцитов в нормальном состоянии, помимо селена, требуется и витамин Е, также обладающий антиоксидантным действием. Например при содержании селена в корме 0,1 мг/кг, хороший рост лососей отмечался, когда количество витамина Е составляло 500 мг .
Низкий уровень витамина Е и наличие в корме полиненасыщенных жиров усиливают влияние дефицита селена. У атлантического лосося это может привести к возникновению болезни "Хитра", или "острой спины", симптомы которой сходны с признаками недостаточности витамина Е и селена. Это заболевание чаще всего встречается при высоких (около 10% и более) концентрациях рыбьего жира в кормах [Watanabe et al., 1997].
По данным Л.Н. Думы [1987], хороший эффект оказывает введение селена в количестве 0,1 мг/кг в комбикорм для сеголеток карпа, где рыбная мука полностью заменена на кормовые дрожжи (паприн), дефицитные по микроэлементам (рецепт ВБС-РЖ-85). Присутствие селена снимает отрицательное влияние этой замены на обмен веществ, резистентность и рост рыб, улучшает физиологическое состояние молоди, повышает ее устойчивость к стрессам и выживаемость в зимний период, а также оказывает благоприятное действие на скорость роста рыб на втором году жизни. Введение в корм селенита натрия (Na2Se3) в количестве 8,6 мг/кг при выращивании в аквакультуре молоди кижуча улучшало ее физиологическое состояние по сравнению с состоянием диких форм.
Большое количество полиненасыщенных жирных кислот в кормах увеличивает потребность рыб в селене и витамине Е. Поэтому целесообразно вводить селен в комбикорма совместно с витамином Е, что позволяет предотвратить токсическое воздействие окисленных жиров на организм рыб.
Йод является составной частью гормонов щитовидной железы (тироксина и трийодтиронина) и тем самым принимает участие в регуляции обмена углеводов, белков и жиров. Он оказывает влияние на развитие и функционирование воспроизводительной системы и рост организма.
Источником йода для рыб являются вода и пища. Потребность рыб в этом элементе по данным Ватанабы с соавторами [T.Watanabe et al, 1997] колеблется от 1 до 4 мг/кг и зависит от концентрации его в воде.
Симптомы недостатка йода проявляются в торможении общего обмена веществ у рыб, что приводит к замедлению роста и снижению устойчивости организма к неблагоприятным воздействиям. У лососевых более 100 лет назад наблюдали гипертрофию щитовидной железы; зоб уменьшался и рассасывался при введении в корм 0,6-1,1 мг/кг йода [Lall, 1989].
Морская вода, растения и животные, живущие в ней, богаты йодом, поэтому морские рыбы не испытывают его дефицита. В пресной воде его концентрация во много раз ниже, чем в морской, и обычно составляет 0,002-0,015 мг/л. Поэтому основное количество йода рыбы должны получать из пищи. Особенно жесткие требования к содержанию йода в корме рыбы предъявляют, если хозяйства располагаются в биогеохимических провинциях, дефицитных по этому элементу (северо-западные и центральные области России, частично Подмосковье, Карелия, многие районы Сибири, Армения, Западная Украина, районы Дальнего Востока и т.д.). В регионах, не испытывающих недостатка йода, потребности рыб удовлетворяются полностью, если в состав комбикормов входит около 7-16% рыбной муки.
Помимо белков рыбной муки, все животные белки, хотя и в меньших количествах, содержат йод. Много йода и в растительном сырье из благополучных по этому элементу провинций.
Благоприятный эффект, выражающийся в стимуляции роста рыб, вызывает введение в состав комбикорма йодистого калия в количестве 1,0-1,5 мг/кг или 2-5% муки из морских водорослей, особенно филлофоры, наиболее богатой йодом, а также обеспечение рыб достаточным количеством естественной пищи [Суховерхов, 1971; Дума, Щербина, 1988].
Медь относится к активным микроэлементам и регулирует многие реакции клеточного дыхания, являясь составной частью ферментов. Недостаток меди приводит к ухудшению работы сердца, мозга, печени, способствует развитию катаракты, угнетает рост. Потребности карпа и форели в меди составляют около 3 мг/кг корма, атлантического лосося — 5 мг/кг (см. табл. 10). У канальных сомов в аквакультуре дефицит меди вызывал пониженную активность в печени перекисной медно-цинковой дисмутазы и цитохром-С-оксидазы сердца. Введение в корм меди в количестве 5 мг/кг предотвращало угнетение ферментов и торможение роста.
Рыбы получают медь из воды и корма. Ее основным депо в организме является печень. По уровню концентрации меди в этом органе судят об обеспеченности рыб этим элементом.
В связи с загрязнением водоемов промышленными стоками содержание меди в воде рыбоводных хозяйств нередко превышает ПДК (0,001 мг/л) в несколько десятков, а иногда и сотен раз (0,01-0,8 мг/л). Кроме того, в наиболее часто употребляемом кормовом сырье ее недостатка для рыб обычно не наблюдается. Скорее всего, беспокойство вызывает ее избыток. По данным И.Н. Остроумовой [2001], наличие меди в корме для теплолюбивого канального сома в количестве 16-32 мг/кг, привело к повышению ее уровня в печени и затормозило рост рыб. В то же время у тепловодного карпа, получавшего повышенные количества меди (30 мг/кг против 4) с высокопитательным гаприном, не отмечали заметных отклонений в росте и физиологическом состоянии за исключением пониженного содержания витамина А в печени. Известно, что медь является антагонистом витамина А и ускоряет процесс его окисления [Орлинский, 1984]. У холодолюбивой форели при повышении дозы меди в рационе до 500-600 мг/кг не обнаружено серьезных отклонений от нормального физиологического состояния даже при дозах свыше 730 мг/кг. Однако были отмечены некроз печени и почек, а также нарушение структуры жаберных лепестков.
По всей вероятности, высокая устойчивость организма рыб к избыточным количествам меди в корме частично связана с малой доступностью для рыб ее различных химических форм (22-32%, по Сергеевой [1989]) и низким содержанием в воде при проведении экспериментов. Влияние избытка меди проявляется еще сильнее при недостатке в корме естественных антиоксидантов (витаминов С, Е). В этом случае медь усиливает перекисное окисление докозапенгаеновой и докозагексаеновой кислот. Это приводит к патологическим изменениям в клеточных мембранах и нарушению функций тканей, например, клеток печени и крови. В последнем случае происходит гемолиз эритроцитов [Kaushik, 1995; Kaushik, Cuzon, 1999].
В коммерческие корма медь вводят обычно в количестве 15-18 мг/кг из-за опасения негативного влияния высоких уровней цинка на абсорбцию меди.
