В количественном отношении второй тип эндогенной экскреции в значительной степени превышает объемы первого.
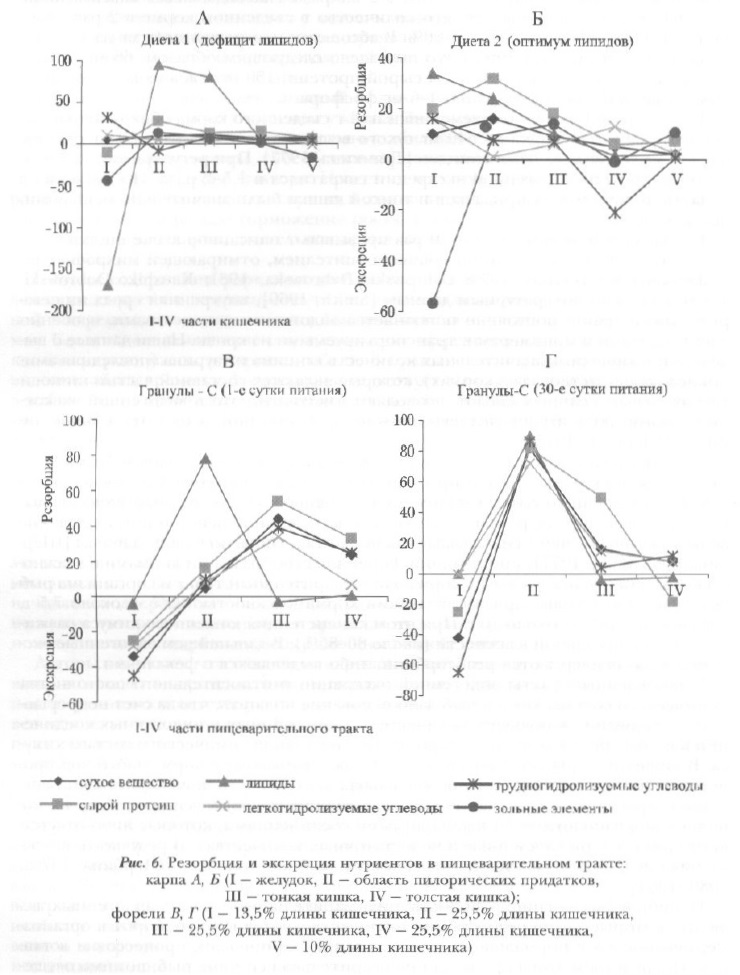
Феномен эндогенной экскреции был обнаружен нами в 70-х годах прошлого столетия как у не имеющего желудка, непрерывно питающегося полифага карпа, так и у порционно питающейся хищной форели, обладающей желудочно-кишечным пищеварением [Щербина, 1973а, 1980; Щербина, Трямкина, 1973; Трямкина, 1977] (рис. 6).
Было установлено, что наибольшая интенсивность экскреции этого типа наблюдается в передних отделах пищеварительного канала. Процесс тесным образом связан с качественным и количественным составом корма и более выражен при резком недостатке или избытке питательных веществ, т.е. в случае несбалансированности питания. Согласно нашим наблюдениям, у карпа при низком уровне белков в диете (<15%) количество выделенного эндогенного азота достигало 25% съеденного.
При наличии в диете менее 2% липидов, их количество в кишечнике было большим, чем поступившее с кормом, в 2-2,5 раза. Наблюдавшееся максимальное выделение кальция превысило его количество в съеденном корме в 2 раза, магния — на 60%, фосфора — на 40-60%. В абсолютном выражении (в мг на 1 г сухого вещества съеденной пищи) это выглядело следующим образом: 66 мг азотсодержащих веществ в пересчете на сырой протеин, 150 мг углеводов, 33 мг липидов, 25 мг кальция, 4 мг магния, 4-6 мг фосфора.
У форели при разовом приеме пищи на 1 г съеденного корма обнаружено выделение в полость желудка до 640 мг сухого вещества, в том числе 160 мг протеина, 140 мг углеводов, 72 мг липидов [Трямкина, 1977]. При регулярном 2-разовом питании уровень эндогенной экскреции сократился в 1,5-2 раза. Поступления в область пилорических придатков и тонкой кишки были значительно меньшими, чем в желудок.
Приведенные величины в 6-20 раз превышают описанное выше поступление эндогенных веществ со слущивающимся эпителием, отмирающей микрофлорой и ферментами [Ogino, 1973; Dabrowski, Dabrowska, 1981; Kaushik, Dobrowski, 1983]. Согласно литературным данным [Smith, 1969], внутренняя среда пищеварительного тракта постоянно пополняется эндогенными веществами, особенно электролитами и мономерами, транспортируемыми из крови. Наши данные о выделении в кишечник значительных количеств глицина и таурина (последняя аминокислота отсутствовала в кормах), которые являются составной частью глико- и таурохолевых желчных кислот, позволяют заключить, что в эндогенной экскреции важную роль играет желчевыделение, особенно при недостатке в пище липидов [Щербина, 1973а].
Значительный объем поступающих в желудок форели углеводов и белков связан с секрецией слизи, продуцируемой железистым аппаратом. Со слизью, желудочным и кишечным соками выделяется основное количество эндогенной воды, которая обеспечивает разжижение химуса до нужной консистенции. Выделение воды тем больше, чем менее увлажнена пища. Так, согласно нашим данным [Щербина, Трямкина, 1977], при питании форели пастообразными кормами с влажностью 45%, уже через час на 1 г корма в пищеварительный тракт из организма рыб поступает 4-6 г воды, при заглатывании корма влажностью 35% — около 7,5 г, влажностью 10% — около 10 г. При этом консистенция кишечного химуса разжижается до нормальной влажности (около 80-85%). В дальнейшем эндогенные экскреты либо подвергаются реабсорбции, либо выделяются с фекалиями.
Установленные факты эндогенной экскреции и относительного постоянства химического состава химуса рыб дали основание полагать, что за счет непрерывного смешивания в больших количествах алиментарных и эндогенных соединений в пищеварительном тракте происходит регуляция химического состава химуса. В кишечнике рыб создаются условия, обеспечивающие нормальный ход процесса пищеварения. Эта приспособительная деятельность пищеварительной системы коррегирует соотношение всасываемых питательных веществ путем их пополнения физиологически необходимыми соединениями, которые либо отсутствуют, либо содержатся в пище в недостаточных количествах. В результате всасываемая смесь приближается к потребностям рыб в нутриентах [Щербина, 1973а, 1980, 1984].
Подобное выделение в полость пищеварительного тракта эндогенных веществ, которые после реабсорбции вновь включаются в кругооборот в организме, способствует нормальному осуществлению химических процессов и в тканях. Посредством этого процесса пищеварительная система рыб, помимо осуществления ранее известных функций, связанных с перевариванием и усвоением пищи (секреция соков, расщепление, всасывание, моторика, выделение), выполняет и обменные функции, направленные на поддержание постоянства концентраций веществ в крови и других средах, составляющих внутреннюю среду организма.
Открытый у рыб феномен эндогенной экскреции позволяет говорить, что та совершенная форма участия пищеварительного аппарата в общем обмене веществ, которая обнаружена у сельскохозяйственных животных [Разенков, 1948] и значительно позже — у человека [Шлыгин, 1974, Гальперин, Лазарев, 1986], достаточно хорошо выражена уже у низших позвоночных. Коррегирующая функция пищеварительной системы четко проявляется уже у карпа, имеющего слабо дифференцированный пищеварительный тракт, вся деятельность которого осуществляется только в кишечнике.
У форели, имеющей более сложный желудочно-кишечный тракт, элементы регуляции выражены отчетливее, чем у карпа. Эволюционно выработанная адаптация форели как хищника к относительно стабильному составу пищи, содержащей все необходимые элементы, способствовала возникновению большей, чем у карпа, зависимости от этого фактора. Известно, что при исключении из диеты форели и лососей незаменимых жирных и аминокислот, а также минералов и витаминов отмечается резкое торможение роста, тогда как у других видов рыб, в частности полифага карпа, длительное время не обнаруживаются симптомы недостаточности.
В случае содержания форели на дисбалансированной диете ее организм немедленно реагирует на изменение качества корма включением механизма эндогенной экскреции, что может вызвать быстрое истощение адаптационных механизмов. Следствие — замедление роста, снижение эффективности использования пищи на прирост массы и нередко гибель. В практическом плане высокая степень эндогенной экскреции у форели свидетельствует о повышенной напряженности в работе всего организма и является показателем несбалансированности рациона, указывая на несоответствие соотношения питательных веществ в корме потребностям рыб.
За последние двадцать лет накоплено определенное количество экспериментальных данных, подтверждающих наличие у высших животных и человека механизма коррегирующей деятельности пищеварительной системы — "питательного гомеостаза".
По трактовке Ю. Гальперина и П. Лазарева [1986], "питательный гомеостаз" — это состояние внутренней среды организма, при котором приспособительные изменения в координированной деятельности физиологических механизмов обеспечивают поддержание в ней концентрации питательных веществ в относительно малом интервале значений, а в пределах этого интервала - на уровне, адекватном внешним и внутренним возмущающим воздействиям."
Авторы отмечают, что опасность нарушения питательного гомеостаза, как правило, возникает в период активного пищеварения. При его затухании все потребности организма удовлетворяются за счет мобилизации ранее ассимилированных веществ из эндогенного депо. В период активного пищеварения гомеостаз может поддерживаться только за счет уравновешивания между процессами всасывания мономеров и скоростями их утилизации, а также депонирования. Он нарушается тогда, когда скорость пополнения внутренней среды всасывающимися веществами, может превысить скорость депонирования той их части, которая не расходуется в данный момент на обеспечение метаболических процессов в тканях. Это превышение приводит к критическому возрастанию концентраций в крови и в других внутренних средах организма всасывающихся веществ. При недостатке каких-либо веществ ответная реакция организма выражается в выделении их из внутренней среды в начальные отделы пищеварительного тракта. Основным и наиболее важным механизмом, обеспечивающим формирование гомеостагированной внутренней среды, и наиболее экономичным способом является постоянная рециркуляция эндогенных веществ.
Наиболее сильное внешнее воздействие на эндогенную экскрецию оказывает степень сбалансированности корма. Только в тех случаях, когда в состав рациона входят все основные питательные вещества, характерные для сбалансированных рационов, эндогенные вещества всасываются уже в начальном отделе пищеварительного тракта. В остальных случаях масса их в химусе возрастает тем значительней, чем ниже исходное содержание этих веществ в рационе. В результате пополнения химуса эндогенной водой и веществами их концентрации стабилизируются в относительно малом (по сравнению с кормом) интервале значений.
Ознакомление с трудом Ю. Гальперина и П. Лазарева [1986], посвященного проблеме пищеварительно-транспортных процессов в кишечнике и разработке теоретических основ энтерального зондового питания человека, еще более убедило нас в универсальности механизма эндогенной экскреции, направленной на поддержание питательного гомеостаза.