По скорости биосинтеза белковой массы бактерии и дрожжи значительно превосходят все известные живые существа. Из дрожжей в основном используются непатогенные виды, относящиеся к родам Candida, Torulopsis, Kluyveromyces, Saccharomyces и др. Культивируемая на различных субстратах биомасса дрожжей обладает целым рядом питательных свойств, полезных для животных. К ним относится способность продуцировать в больших количествах полноценные по аминокислотном)' составу белки, синтезировать витамины, а также иметь в своем составе углеводы, достаточно хорошо доступные рыбам, и ряд других веществ.
По набору и соотношению незаменимых аминокислот (за исключением метионина) их белки способны удовлетворять потребности основных видов культивируемых рыб. Однако дрожжи в своем составе содержат много небелкового азота. Общая сумма безводных остатков аминокислот существенно ниже показателя "сырого протеина" (общий азот х коэффициент 6,25). Н. Крылова с сотрудниками [1983] установили, что зависимость между содержанием общего азота и белка в биомассе дрожжей имеет линейный характер и описывается уравнением
белок = общий азот х 5,18 + 2,64, где 5,18 — коэффициент.
Однако в справочных таблицах по химическому составу сырья (в том числе и приведенных в разделе) это обстоятельство не учтено, и сырой протеин рассчитан путем умножения общего азота на коэффициент 6,25.
Как указано выше, отличительная особенность сырого протеина дрожжей — это присутствие значительных количеств небелковых азотистых соединений (до 25% по массе и 50% по отношению к сырому протеину). Большая часть его приходится на нуклеиновые кислоты, в частности РНК (до 15-20% сухого вещества). Известно, чем выше содержание общего азота, тем больше РНК в биомассе дрожжей. Диапазон их содержания в дрожжевых продуктах в среднем составляет 8-10%, в молодых культурах —до 15-20%, в бактериальной массе — 12-14% [Крылова и др., 1983; Остроумова, 1991; Судакова, 1998]. Это ограничивает применение дрожжей в кормлении сельскохозяйственных животных, т.к. они не обладают способностью к достаточно полному расщеплению единовременного поступающего с дрожжами большого количества РНК. Недоокисленные продукты распада РНК оказывают токсическое действие, в результате которого возникают разнообразные патологические изменения в организме животных и птиц. Рыбы, в отличие от наземных позвоночных, не накапливают продуктов нуклеинового обмена. Они способны расщеплять их до легкорастворимых соединений и либо утилизировать в процессах обмена, либо удалять из организма через жабры и почки в виде аммиачных солей и мочевины. Это объяснимо, если принять во внимание, что в естественной пище рыб — зоопланктоне и бентосе, содержание РНК также может достигать 5-10% [Остроумова, 2001].
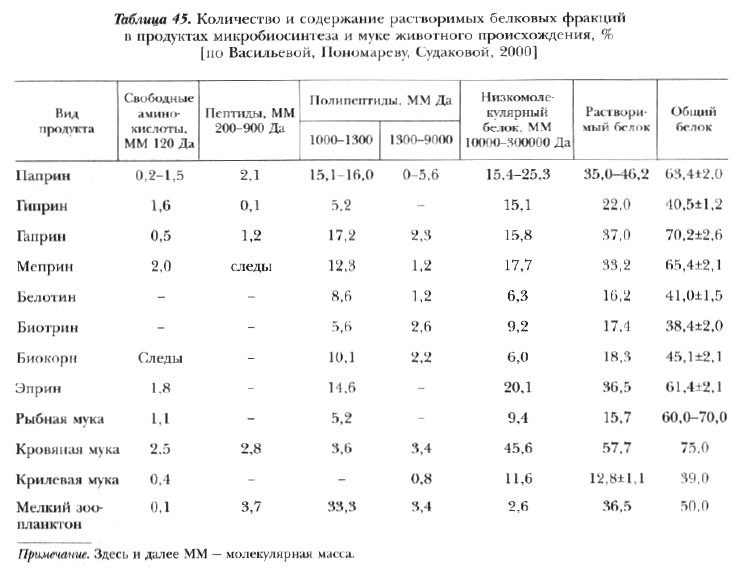
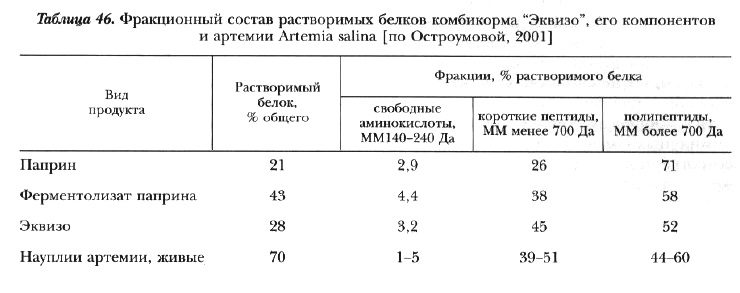
Важное достоинство некоторых видов дрожжей (например, эприна) и микробной биомассы (гаприна)-наличие большого количества растворимых белков, имеющих в своем составе промежуточные продукты белкового синтеза: свободные аминокислоты, пептиды, низкомолекулярные полипептиды, что свойственно очень быстро размножающимся и быстрорастущим организмам (табл. 45 и 46).
По этому признаку физико-химический состав их белков сближается с таковым белков мелких форм водных беспозвоночных, которые служат естественной пищей для молоди большинства рыб. Последняя особенность послужила основанием для использования продуктов микробиосинтеза в качестве основных компонентов в стартовых кормах для некоторых видов рыб. Личинки этих рыб имеют очень короткий период развития и при переходе к активному питанию еще не имеют достаточно развитой пищеварительной системы. Присутствие в корме мелких фракций белка, не требующих глубокой ферментативной обработки и легко усвояемых в нативном виде, обеспечивает потребности личинок в структурном материале для роста еще до начала нормального функционирования пищеварительных желез и других дефинитивных органов [Остроумова и др., 1979; Остроумова, Дементьева, 1981; Остроумова, Ильина, 1981; Канидьев и др., 1983; Остроумова, 1983а, 2001].
Другое достоинство ряда дрожжей, повышающее их ценность в питании ранней молоди рыб, — высокая протеазная активность, которая проявляется при pH 1,6 и 7,2, т.е. в среде, свойственной как безжелудочным рыбам, так и рыбам с желудочно-кишечным пищеварением. Она обусловлена присутствием в дрожжевых клетках глютатиона-трипептида, представляющего собой соединение глицина, цистеина и глютаминовой кислоты. Глютатион физиологически активен и оказывает влияние на активность многих ферментов, связанных с превращением белков, участвует в связывании кислорода гемоглобином [Чернышов, Панин, 2000].
К отрицательных свойствам ряда дрожжей относятся большая толщина и высокая прочность клеточных оболочек, делающие их содержимое плохо доступным для пищеварительных ферментов животных. Это тем более существенно, что в биомассе дрожжей они могут составлять 20-30% и более. Поэтому в целях повышения доступности для организма животных питательных веществ дрожжей используются специальные технологические приемы. В их число входит разрушение дрожжевых клеток дроблением, а так же их внутриклеточного содержимого посредством гидролиза. При разрушении клеточной оболочки состав белков и других веществ не изменяется.
При гидролизе под действием химических реагентов (кислот, щелочей) или вводимых ферментов происходит расщепление высокомолекулярных соединений до низкомолекулярных. В частности, труднопереваримые полисахариды оболочек дрожжей подвергаются деструкции до легко доступных фрагментов. Ферментативный гидролиз позволяет получать в физиологически активной форме до 94% всех аминокислот. Биологически и экономически наиболее эффективен автолиз, который осуществляется с помощью собственных активированных протеолитических ферментов клетки [Попова и др., 1988].
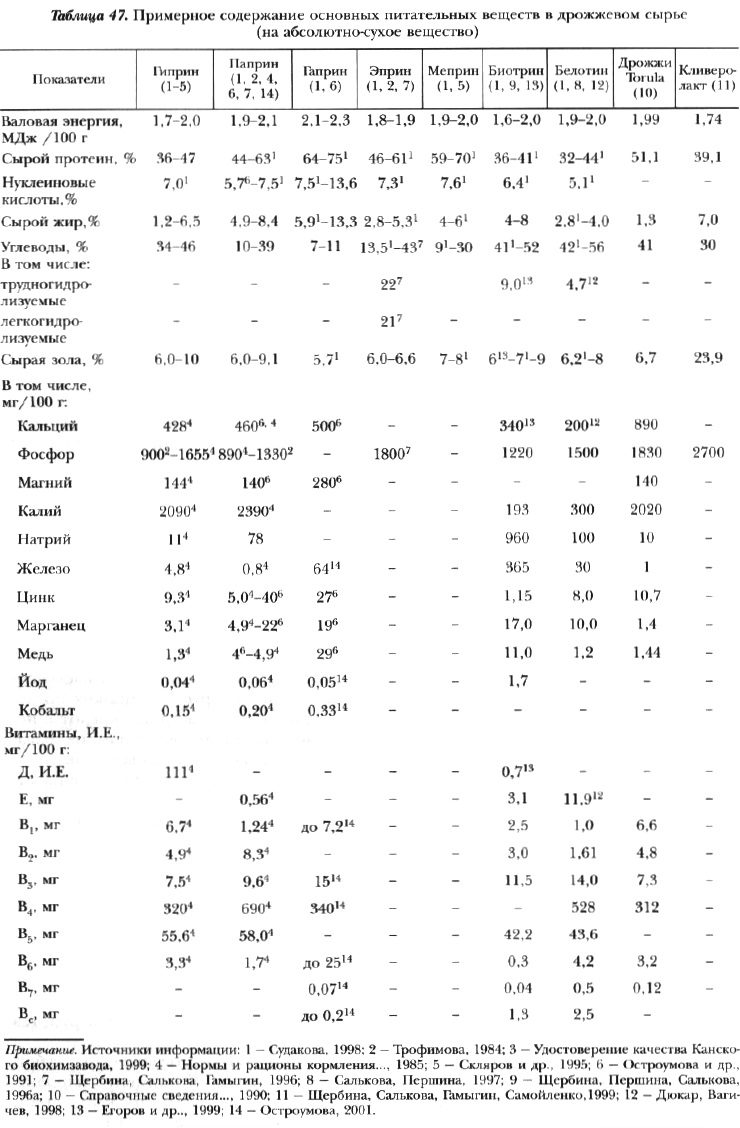
По уровню витаминов группы В дрожжи значительно превосходят все виды кормового сырья, причем эти витамины усваиваются в животном организме значительно лучше, чем синтетические. Благодаря высокому' содержанию белка и витаминов (табл. 47) некоторые виды кормовых дрожжей получили название белково-витаминных концентратов — БВК.
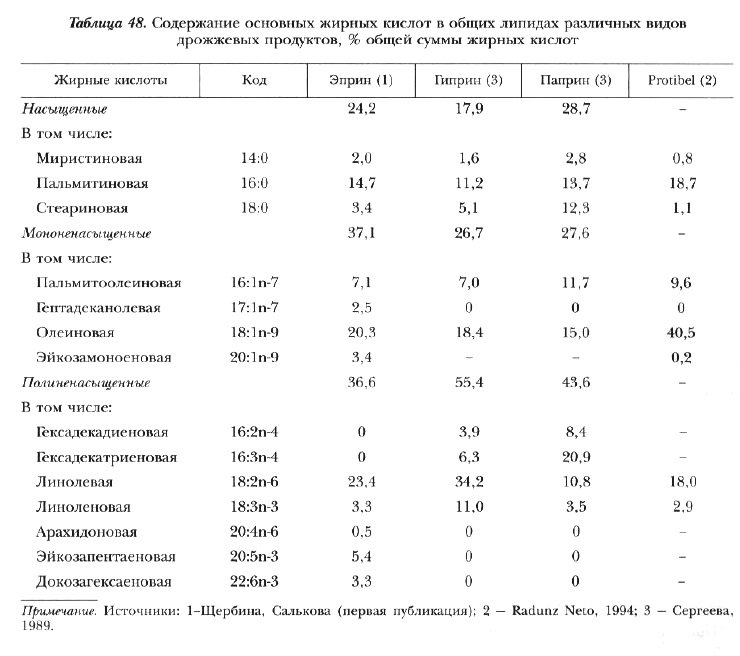
В состав липидов дрожжей входят фосфолипиды, триглицериды, стерины, воска и жирные кислоты. Соотношение поли-, мононенасыщенных и насыщенных жирных кислот по отдельным видам дрожжей варьирует в широких пределах (табл. 48). Отличительная особенность — присутствие жирных кислот с нечетным числом углеродных атомов, которые, по некоторым сведениям, оказывают неблагоприятное воздействие на животных. Однако позднее было показано, что в малых количествах они присутствуют в липидах водных беспозвоночных и рыб [Касаткина, 1988; Сергеева, 1989; Radunz, Neto, 1994].
Изучение у рыб метаболизма выявило их позитивную роль в регуляции содержания фосфолипидов и холестерина в плазме крови, а также в мембранных структурах клеток [Аствацагурьян, 1989].
Селекционированы специальные штаммы дрожжей (например, Hansenula polymorpha), которые продуцируют в большом количестве липиды, в частности незаменимую для тепловодных рыб арахидоновую кислоту [Дедюхина и др., 1980].
Для производства микробиологической продукции в основном используют углеводное сырье, которое обычно не применяется в кормлении животных (отходы пищевого и деревообрабатывающих производств, бумажной промышленности) и углеводороды- отходы нефте- и газопереработки (н-парафины нефти, газ метан и др.).
Получаемые белковые продукты имеют название, соответствующее виду субстратов, на которых ведется культивация микроорганизмов. К примеру: гидролизные дрожжи-гиприн, алкановые дрожжи-паприн, этанольные дрожжи-эприн, метанольные дрожжи-меприн, метанокислящие бактерии-гаприн и т.д.
В упрощенном виде технология кормовых дрожжей согласно Н. Калунянцу и его соавторам [1980], включает следующие процессы: 1) подготовка субстрата культивирования; 2) приготовление оптимизирующих добавок (обычно аммиачной воды и растворов солей сернокислого аммония, диаммонийфосфата, суперфосфата, сернокислого и хлористого магния и калия, углекислого кальция, орто-фосфорной кислоты и др.); 3) подготовка питательной среды; 4) культивирование посевного материала; 5) выращивание биомассы дрожжей в "ферментерах" при определенной температуре, pH и концентрации газов; 6) отделение биомассы от культурального материала путем одно— двукратного сепарирования, промывка ее водой; 7) концентрирование и плазмолиз (автолиз) дрожжей в многоступенчатой вакуумвыпаривательной установке; 8) сушка в распылительных сушилках; 9) расфасовка и упаковка готового продукта.
В зависимости от вида дрожжей они могут вводиться в продукционные и стартовые комбикорма для различных видов рыб в количествах 2-62% для хищных рыб и 5-56% — для всеядных и растительноядных [Jauncey, 1995].
Отрицательным свойством дрожжевых продуктов может быть наличие в них живых клеток продуцента, которые вызывают у рыб различные микозы. В частности, кандидамикоз характеризуется не только поражением слизистых оболочек пищеварительного тракта, но и жабр с образованием на них гифов грибов. Болезнь также возникает при применении плохо хранившихся комбикормов и поражает прежде всего ослабленных рыб.
В настоящее время в нашей стране прекращен выпуск продуктов микробиосинтеза, обладающих наиболее высокой питательностью для рыб (эприна, паприна, гаприна, их ферментолизатов). Это обстоятельство наряду с дефицитом высококачественного белкового сырья резко осложняет производство полноценных комбикормов, особенно для индустриальных хозяйств, и создает большие проблемы при разработке и выпуске эффективных стартовых комбикормов для личинок карповых, сиговых и осетровых рыб.